Stanovení respiračního koeficientu (RK) rostlin. Stanovení respiračního koeficientu Respirační koeficient jeho hodnota
respirační koeficient (RK)
poměr objemu oxidu uhličitého uvolněného plícemi k objemu kyslíku absorbovaného za stejnou dobu; hodnota D.c., když je subjekt v klidu, závisí na druhu potravinových látek oxidovaných v těle.
Encyklopedický slovník, 1998
dýchacího kvocientu
poměr objemu oxidu uhličitého uvolněného při dýchání za určitou dobu k objemu kyslíku absorbovaného za stejnou dobu. Charakterizuje rysy výměny plynů a metabolismu u živočichů a rostlin. U zdravého člověka je to přibližně 0,85.
Respirační koeficient
poměr objemu oxidu uhličitého uvolněného z těla k objemu kyslíku absorbovaného za stejnou dobu. Označuje:
Stanovení DC je důležité pro studium charakteristik výměny plynů a metabolismu u zvířat a rostlinných organismů. Když jsou sacharidy v těle oxidovány a kyslík je plně dostupný, DC je 1, tuky ≈ 0,7, bílkoviny ≈ 0,8. U zdravého člověka v klidu je DC 0,85 ╠ 0,1; při mírné práci, stejně jako u zvířat, která se živí převážně rostlinnou potravou, se blíží 1. U člověka při velmi dlouhé práci, hladovění, u masožravců (predátorů), stejně jako při zimním spánku, kdy kvůli omezeným zásobám sacharidů v těle, disimilace zvyšuje tuk, DC je asi 0,7. DC přesahuje 1 při intenzivním ukládání tuků v těle tvořených ze sacharidů dodávaných s potravou (např. u lidí při obnově normální hmotnosti po hladovění, po dlouhodobých nemocech, stejně jako u zvířat při výkrmu). DC se zvyšuje na 2 při zvýšené práci a hyperventilaci plic, kdy se z těla uvolňuje další CO2, který byl ve vázaném stavu. Ještě vyšších hodnot DC dosahuje u anaerobů, ve kterých většina uvolněného CO2 vzniká bezkyslíkatou oxidací (fermentací). DK pod 0,7 se vyskytuje u onemocnění spojených s poruchami metabolismu, po těžké fyzické práci.
U rostlin závisí DK na chemické povaze dýchacího substrátu, obsahu CO2 a O2 v atmosféře a dalších faktorech, charakterizuje tak specifika a podmínky dýchání. Když buňka používá k dýchání sacharidy (sazenice obilnin), DC je přibližně 1, tuky a bílkoviny (klíčící olejná semena a luštěniny) ≈ 0,4≈0,7. Při nedostatku O2 a obtížném přístupu (semena s tvrdou skořápkou) je DC 2≈3 nebo více; vysoký DC je také charakteristický pro buňky růstového bodu.
(DC) je poměr objemu oxidu uhličitého uvolněného při dýchání k objemu absorbovaného kyslíku.
Hodnota dýchacího koeficientu rostliny
DC hodnota označuje jak povahu materiálu oxidovaného během dýchání, tak typ dýchání; může být rovna, větší nebo menší než jedna. Při oxidaci sacharidů se objemy vyměněných plynů oxidu uhličitého a kyslíku rovnají a poměr C0 2: 0 2 je roven jednotce. V tomto případě se kyslík spotřebovaný při dýchání využívá pouze k oxidaci uhlíku na oxid uhličitý, protože poměr vodíku a kyslíku v molekule glukózy je takový, že pro oxidaci vodíku na vodu je v cukru dostatek kyslíku. samotná molekula. Při oxidaci řady organických kyselin je respirační koeficient rostlin větší než jedna. Kyselina šťavelová je tedy sloučenina bohatší na kyslík než sacharidy. Kyslík přítomný v molekule nestačí pouze k oxidaci vodíku na vodu, ale část z něj zůstává na oxidaci uhlíku; proto pro úplnou oxidaci dvou molekul kyseliny šťavelové stačí jedna molekula kyslíku: 2C 2 H 2 O 4 + O 2 → 4CO 2 + 2H 2 O, DC (4CO 2: O 2) je v tomto případě rovno až 4. V případech, kdy rostlina dýchá díky bílkovinám nebo tukům, jejichž molekula obsahuje hodně vodíku a uhlíku a málo kyslíku je menší než jedna, protože za účelem oxidace veškerého uhlíku a vodíku, které se v těchto sloučeninách nacházejí; , je nutné absorbovat velké množství kyslíku. Když je kyselina stearová oxidována, oxidační reakce bude probíhat následovně: C18H26O2 + 2602 → 18CO2 + 18H20. DC (18CO2:2602) je 0,69. V případě oxidace sacharidů se tedy DC rovná jedné, organické kyseliny - více než jedna, bílkoviny a tuky - méně než jedna.
Tepelný efekt při dýchání rostlin
Tepelný efekt bude mít opačnou hodnotu DC: maximální tepelný efekt bude při oxidaci tuků, protože se jedná o nejvíce redukované sloučeniny. Závislost hodnoty DC na charakteru dýchacího materiálu je pozorována pouze při dostatku kyslíku v prostředí a rostlinných pletivech. Nicméně během oxidace stejného dýchacího materiálu, ale s nedostatkem kyslíku v prostředí a rostlinných tkáních, se mohou hodnoty DC také změnit. Pokud je kyslíku málo, pak oxidace neproběhne do konce a kromě oxidu uhličitého a vody vznikají organické kyseliny, které jsou oxidovanější než sacharidy. V tomto případě bude DC menší než jedna, protože část absorbovaného kyslíku zůstane v molekulách vytvořených organických kyselin, zatímco se uvolní méně oxidu uhličitého. Uvolní se méně energie, protože část bude uložena v organických kyselinách.1. Jaký proces zajišťuje uvolňování energie v těle? Jaká je její podstata?
Disimilace (katabolismus), tj. rozklad buněčných struktur a sloučenin těla s uvolňováním energie a produktů rozpadu.
2. Jaké živiny dodávají tělu energii?
Sacharidy, tuky a bílkoviny.
3. Vyjmenujte hlavní metody stanovení množství energie ve vzorku výrobku.
Fyzikální kalorimetrie; fyzikálně-chemické metody stanovení množství živin ve vzorku s následným výpočtem energie v něm obsažené; podle tabulek.
4. Popište podstatu metody fyzikální kalorimetrie.
Vzorek produktu se spálí v kalorimetru a následně se vypočítá uvolněná energie na základě stupně zahřátí vody a materiálu kalorimetru.
5. Napište vzorec pro výpočet množství tepla uvolněného při spalování produktu v kalorimetru. Dešifrujte jeho symboly.
Q = MvSv (t 2 - t 1) + MkSk (t 2 - t 1) - Q®,
kde Q je množství tepla, M je hmotnost (w - voda, k - kalorimetr), (t 2 - t 1) je teplotní rozdíl mezi vodou a kalorimetrem po a před spálením vzorku, C je měrné teplo kapacita, Qo je množství tepla generovaného okysličovadlem.
6. Jaké jsou fyzikální a fyziologické kalorické koeficienty živiny?
Množství tepla uvolněného při spalování 1 g látky v kalorimetru a v těle, resp.
7. Kolik tepla se uvolní, když se v kalorimetru spálí 1 g bílkovin, tuků a sacharidů?
1g bílkovin – 5,85 kcal (24,6 kJ), 1g tuku – 9,3 kcal (38,9 kJ), 1g sacharidů – 4,1 kcal (17,2 kJ).
8. Formulujte Hessův termodynamický zákon, na jehož základě se vypočítá energie vstupující do těla na základě množství natrávených bílkovin, tuků a sacharidů.
Termodynamický efekt závisí pouze na obsahu tepla v počátečních a konečných reakčních produktech a nezávisí na přechodných přeměnách těchto látek.
9. Kolik tepla se uvolní při oxidaci 1 g bílkovin, 1 g tuků a 1 g sacharidů v těle?
1 g bílkovin – 4,1 kcal (17,2 kJ), 1 g tuku – 9,3 kcal (38,9 kJ), 1 g sacharidů – 4,1 kcal (17,2 kJ).
10. Vysvětlete důvod rozdílu mezi fyzikálním a fyziologickým kalorickým koeficientem pro bílkoviny. V jakém případě je větší?
V kalorimetru (fyzikálním koeficientu) se bílkovina rozkládá na konečné produkty - CO 2, H 2 O a NH 3 za uvolnění veškeré energie v nich obsažené. V těle (fyziologický koeficient) se bílkoviny rozkládají na CO 2, H 2 O, močovinu a další látky metabolismu bílkovin, které obsahují energii a jsou vylučovány močí.
Stanoví se obsah bílkovin, tuků a sacharidů v potravinářských výrobcích, jejich množství se vynásobí odpovídajícími fyziologickými kalorickými koeficienty, sečte se a od součtu se odečte 10 %, které se nevstřebávají v trávicím traktu (ztráty stolicí).
12. Vypočítejte (v kcal a kJ) energetický příjem při příjmu 10 g bílkovin, tuků a sacharidů do těla potravou.
Q = 4,110 + 9,310 + 4,110 = 175 kcal. (175 kcal - 17,5 kcal) x 4,2 kJ, kde 17,5 kcal je energie nestrávených živin (ztráty stolicí - asi 10 %). Celkem: 157,5 kcal (661,5 kJ).
Kalorimetrie: přímá (metoda Atwater-Benedict); nepřímé, nebo nepřímé (metody Krogh, Shaternikov, Douglas - Holden).
14. Na čem je založen princip přímé kalorimetrie?
Na přímém měření množství tepla generovaného tělem.
15. Stručně popište konstrukci a princip fungování kamery Atwater-Benedict.
Komora, ve které je testovaný subjekt, je tepelně izolována od okolí, její stěny neabsorbují teplo, jsou to radiátory, kterými proudí voda. Na základě stupně zahřátí určitého množství vody se vypočítá množství tepla spotřebovaného tělem.
16. Na čem je založen princip nepřímé (nepřímé) kalorimetrie?
Výpočtem množství uvolněné energie podle údajů o výměně plynu (absorbovaný O 2 a uvolněný CO 2 za den).
17. Proč lze množství energie uvolněné tělem vypočítat na základě směnných kurzů plynů?
Protože množství O 2 spotřebovaného tělem a uvolněného CO 2 přesně odpovídá množství zoxidovaných bílkovin, tuků a sacharidů, a tedy i energií spotřebované tělem.
18. Jaké koeficienty se používají pro výpočet spotřeby energie nepřímou kalorimetrií?
Respirační koeficient a kalorický ekvivalent kyslíku.
19. Co se nazývá respirační koeficient?
Poměr objemu oxidu uhličitého uvolněného tělem k objemu kyslíku spotřebovaného za stejnou dobu.
20. Vypočítejte respirační koeficient (RC), pokud je známo, že vdechovaný vzduch obsahuje 17 % kyslíku a 4 % oxidu uhličitého.
Protože atmosférický vzduch obsahuje 21 % O 2, je procento absorbovaného kyslíku 21 % - 17 %, tj. 4 %. CO 2 ve vydechovaném vzduchu je také 4 %. Odtud
21. Na čem závisí dechový koeficient?
22. Jaký je respirační koeficient při oxidaci v těle na konečné produkty bílkovin, tuků a sacharidů?
Při oxidaci bílkovin – 0,8, tuků – 0,7, sacharidů – 1,0.
23. Proč je respirační kvocient nižší u tuků a bílkovin než u sacharidů?
Více O2 se spotřebuje na oxidaci bílkovin a tuků, protože obsahují méně intramolekulárního kyslíku než sacharidy.
24. K jaké hodnotě se přibližuje dechový kvocient člověka na začátku intenzivní fyzické práce? Proč?
Do jedné, protože zdrojem energie jsou v tomto případě především sacharidy.
25. Proč je dechový koeficient člověka v prvních minutách po intenzivní a dlouhodobé fyzické práci větší než jedna?
Protože se uvolňuje více CO 2, než se spotřebuje O 2, protože kyselina mléčná nahromaděná ve svalech vstupuje do krve a vytlačuje CO 2 z bikarbonátů.
26. Jak se nazývá kalorický ekvivalent kyslíku?
Množství tepla uvolněného tělem při konzumaci 1 litru O2.
27. Na čem závisí kalorický ekvivalent kyslíku?
Z poměru bílkovin, tuků a sacharidů zoxidovaných v těle.
28. Jaký je kalorický ekvivalent kyslíku při oxidaci v těle (při procesu disimilace) bílkovin, tuků a sacharidů?
Pro bílkoviny - 4,48 kcal (18,8 kJ), pro tuky - 4,69 kcal (19,6 kJ), pro sacharidy - 5,05 kcal (21,1 kJ).
29. Stručně popište proces stanovení spotřeby energie metodou Douglas-Holden (úplná analýza plynu).
Během několika minut subjekt vdechuje atmosférický vzduch a vydechovaný vzduch je shromažďován do speciálního vaku, jeho množství je měřeno a je provedena analýza plynů ke stanovení objemu spotřebovaného kyslíku a uvolněného CO 2 . Vypočítá se respirační koeficient, pomocí kterého se z tabulky zjistí odpovídající kalorický ekvivalent O 2, který se následně vynásobí objemem spotřebovaného O 2 za dané časové období.
30. Stručně popište metodu M. N. Shaternikova pro stanovení energetického výdeje u zvířat v experimentu.
Zvíře se umístí do komory, do které se při spotřebování přivádí kyslík. CO 2 uvolněný při dýchání je absorbován alkáliemi. Uvolněná energie se vypočítá na základě množství spotřebovaného O2 a průměrného kalorického ekvivalentu O2: 4,9 kcal (20,6 kJ).
31. Vypočítejte spotřebu energie za 1 minutu, pokud je známo, že subjekt spotřeboval 300 ml O 2 . Respirační koeficient je 1,0.
DK = 1,0, odpovídá kalorickému ekvivalentu kyslíku rovnému 5,05 kcal (21,12 kJ). Proto spotřeba energie za minutu = 5,05 kcal x 0,3 = 1,5 kcal (6,3 kJ).
32. Stručně popište proces stanovení spotřeby energie pomocí Kroghovy metody u člověka (neúplná analýza plynů).
Subjekt inhaluje kyslík z metabolimetrického vaku, vydechovaný vzduch se vrací do stejného vaku, který předtím prošel absorbérem CO2. Na základě údajů z metabolimetru se určí spotřeba O2 a vynásobí se kalorickým ekvivalentem kyslíku 4,86 kcal (20,36 kJ).
33. Vyjmenujte hlavní rozdíly ve výpočtu spotřeby energie metodou Douglas-Holden a Krogh.
Metoda Douglas–Holden zahrnuje výpočet spotřeby energie na základě údajů z kompletní analýzy plynu; Kroghova metoda - pouze objemem spotřebovaného kyslíku pomocí kalorického ekvivalentu kyslíku charakteristického pro bazální metabolické poměry.
34. Co se nazývá bazální metabolismus?
Minimální spotřeba energie, která zajišťuje homeostázu za standardních podmínek: v bdělém stavu, s maximálním svalovým a emočním odpočinkem, nalačno (12 - 16 hodin bez jídla), při příjemné teplotě (18 - 20 C).
35. Proč se bazální metabolismus stanovuje za standardních podmínek: maximální svalový a emoční klid, nalačno, při příjemné teplotě?
Protože fyzická aktivita, emoční stres, příjem potravy a změny okolní teploty zvyšují intenzitu metabolických procesů v těle (spotřebu energie).
36. Jaké procesy spotřebovávají bazální metabolickou energii v těle?
K zajištění životních funkcí všech orgánů a tkání těla, buněčné syntézy a udržení tělesné teploty.
37. Jaké faktory určují hodnotu správného (průměrného) bazálního metabolismu zdravého člověka?
Pohlaví, věk, výška a tělesná hmotnost (váha).
38. Jaké faktory kromě pohlaví, hmotnosti, výšky a věku určují hodnotu skutečného (skutečného) bazálního metabolismu zdravého člověka?
Životní podmínky, kterým je tělo přizpůsobeno: trvalý pobyt v chladném klimatickém pásmu zvyšuje bazální metabolismus; dlouhodobá vegetariánská strava – redukuje.
39. Vyjmenujte způsoby, jak zjistit množství správného bazálního metabolismu u člověka. Jaká metoda se používá k určení hodnoty skutečného bazálního metabolismu člověka v praktické medicíně?
Podle tabulek, podle vzorců, podle nomogramů. Kroghova metoda (neúplná analýza plynů).
40. Jaká je hodnota bazálního metabolismu u mužů a žen za den a také na 1 kg tělesné hmotnosti za den?
Pro muže 1500 – 1700 kcal (6300 – 7140 kJ), nebo 21 – 24 kcal (88 – 101 kJ)/kg/den. Ženy mají přibližně o 10 % méně než tato hodnota.
41. Je bazální metabolismus přepočtený na 1 m 2 tělesného povrchu a na 1 kg tělesné hmotnosti u teplokrevníků a lidí stejný?
Při přepočtu na 1 m 2 tělesného povrchu u teplokrevníků různých druhů a lidí jsou ukazatele přibližně stejné, při přepočtu na 1 kg hmotnosti jsou velmi odlišné.
42. Co se nazývá pracovní burza?
Kombinace bazálního metabolismu a dodatečného energetického výdeje, který zajišťuje fungování organismu v různých podmínkách.
43. Uveďte faktory, které zvyšují spotřebu energie tělem. Co se nazývá specifický dynamický účinek jídla?
Fyzická a psychická zátěž, emoční stres, změny teplot a dalších podmínek prostředí, specifické dynamické účinky potravy (zvýšená spotřeba energie po jídle).
44. O kolik procent se spotřeba energie v těle zvýší po konzumaci bílkovin a smíšených potravin, tuků a sacharidů?
Po konzumaci bílkovinných potravin - o 20 - 30%, mixovaných potravin - o 10 - 12%.
45. Jak okolní teplota ovlivňuje energetický výdej těla?
Teplotní změny v rozmezí 15 – 30 C výrazně neovlivňují energetickou spotřebu organismu. Při teplotách pod 15C a nad 30C se spotřeba energie zvyšuje.
46.Jak se mění metabolismus při okolní teplotě pod 15? co na tom záleží?
Rostoucí. Tím se zabrání prochladnutí těla.
47. Jak se nazývá výkonnost těla při svalové práci?
Vyjádřeno v procentech, poměr energie ekvivalentní užitečné mechanické práci k celkové energii vynaložené na provedení této práce.
48. Uveďte vzorec pro výpočet koeficientu výkonu (efektivnosti) u člověka při svalové práci, uveďte jeho průměrnou hodnotu, dešifrujte prvky vzorce.

kde A je energie ekvivalentní užitečné práci, C je celková spotřeba energie, e je spotřeba energie za stejnou dobu v klidu. Účinnost je 20 %.
49. Kterým živočichům se říká poikilotermní a homeotermní?Poikilotermní živočichové (studenokrevní) - s nestabilní tělesnou teplotou v závislosti na teplotě okolí; homeotermní (teplokrevní) - živočichové se stálou tělesnou teplotou nezávislou na okolní teplotě.
50. Jaký význam má pro tělo stálost tělesné teploty? Ve kterých orgánech probíhá proces tvorby tepla nejintenzivněji?
Poskytuje vysokou úroveň vitální aktivity relativně bez ohledu na okolní teplotu. Ve svalech, plicích, játrech, ledvinách.
51. Vyjmenujte druhy termoregulace. Formulujte podstatu každého z nich.
Chemická termoregulace - regulace tělesné teploty změnou intenzity produkce tepla; fyzikální termoregulace – změnou intenzity přenosu tepla.
52. Jaké procesy zajišťují přenos tepla?
Tepelné záření (sálání), odpařování tepla, vedení tepla, konvekce.
53. Jak se mění průsvit krevních cév kůže, když okolní teplota klesá a stoupá? Jaký je biologický význam tohoto jevu?
Při poklesu teploty se krevní cévy v kůži zužují. Se stoupající okolní teplotou se krevní cévy v kůži rozšiřují. Faktem je, že změna šířky lumenu krevních cév, regulace přenosu tepla, pomáhá udržovat konstantní tělesnou teplotu.
54. Jak a proč se při silné stimulaci sympatoadrenálního systému mění tvorba a přenos tepla?
Produkce tepla se zvýší stimulací oxidačních procesů a přenos tepla se sníží v důsledku zúžení kožních cév.
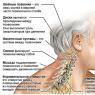
55. Vyjmenujte oblasti lokalizace termoreceptorů.
Kůže, kožní a podkožní cévy, vnitřní orgány, centrální nervový systém.
56. V jakých částech a strukturách centrálního nervového systému se nacházejí termoreceptory?
V hypotalamu retikulární formace středního mozku, v míše.
57. Ve kterých částech centrálního nervového systému se nacházejí centra termoregulace? Která struktura centrálního nervového systému je nejvyšším centrem termoregulace?
V hypotalamu a míše. Hypotalamus.
58. Jaké změny nastanou v těle při dlouhodobé absenci tuků a sacharidů ve stravě, ale při optimálním příjmu bílkovin z potravy (80 - 100 g denně)? Proč?
Dojde k nadměrné spotřebě dusíku v těle nad jeho příjmem a úbytkem hmotnosti, protože náklady na energii budou hrazeny převážně z bílkovin a tukových zásob, které se nedoplní.
59. V jakém množství a v jakém poměru by měly být bílkoviny, tuky a sacharidy obsaženy ve stravě dospělého člověka (průměrná verze)?
Bílkoviny – 90 g, tuky – 110 g, sacharidy – 410 g, poměr 1:1, 2:4, 6.
60. Jak se mění stav těla s nadměrným příjmem tuku?
Rozvíjí se (předčasně) obezita a ateroskleróza. Obezita je rizikovým faktorem pro rozvoj kardiovaskulárních onemocnění a jejich komplikací (infarkt myokardu, cévní mozková příhoda aj.) a zkrácení délky života.
1. Jaký je poměr bazálních rychlostí metabolismu u dětí prvních 3–4 let života, během puberty, ve věku 18–20 let a dospělých (kcal/kg/den)?
Do 3–4 let mají děti přibližně 2x více, v pubertě – 1,5x více než dospělí. Ve věku 18–20 let odpovídá normě dospělých.
2. Nakreslete graf změn bazálního metabolismu u chlapců s věkem (u dívek je bazální metabolismus o 5 % nižší).

3. Čím se vysvětluje vysoká intenzita oxidačních procesů u dítěte?
Vyšší úroveň metabolismu mladých tkání, relativně velký povrch těla a samozřejmě větší výdej energie na udržení stálé tělesné teploty, zvýšená sekrece hormonů štítné žlázy a norepinefrinu.
4. Jak se mění náklady na energii pro růst v závislosti na věku dítěte: do 3 měsíců života, před nástupem puberty, během puberty?
V prvních 3 měsících po narození se zvyšují, pak postupně klesají a během puberty se opět zvyšují.
5. Z čeho se skládá celkový energetický výdej 1letého dítěte a jak je procentuálně rozdělen ve srovnání s dospělým?
U dítěte: 70 % připadá na bazální metabolismus, 20 % na pohyb a udržení svalového tonusu, 10 % na specifický dynamický účinek potravy. U dospělého: 50 – 40 – 10 %, resp.
6. Vydávají dospělí nebo děti ve věku 3–5 let více energie při provádění svalové práce, aby dosáhli stejného užitečného výsledku, kolikrát a proč?
Děti 3 až 5krát, protože mají méně dokonalou koordinaci, což vede k nadměrným pohybům, což má za následek výrazně méně užitečnou práci pro děti.
7. Jak se mění energetický výdej, když dítě pláče, o kolik procent a v důsledku čeho?
Zvyšuje se o 100 - 200 % v důsledku zvýšené produkce tepla v důsledku emočního vzrušení a zvýšené svalové aktivity.
8. Jakou část (v procentech) energetického výdeje dítěte zajišťují bílkoviny, tuky a sacharidy? (srovnej s normou pro dospělé).
Kvůli bílkovinám - 10%, kvůli tukům - 50%, kvůli sacharidům - 40%. U dospělých – 20 – 30 – 50 %, resp.
9. Proč se děti, zvláště v kojeneckém věku, rychle přehřívají, když se zvýší okolní teplota? Snášejí děti snáze zvýšení nebo snížení okolní teploty?
Protože děti mají zvýšenou produkci tepla, nedostatečné pocení a tedy i odpařování tepla, je nezralé centrum termoregulace. Degradace.
10. Pojmenujte bezprostřední příčinu a vysvětlete mechanismus rychlého ochlazení dětí (zejména kojenců) při poklesu okolní teploty.
Zvýšený přenos tepla u dětí v důsledku poměrně velkého tělesného povrchu, bohatého prokrvení pokožky, nedostatečné tepelné izolace (tenká kůže, nedostatek podkožního tuku) a nezralosti termoregulačního centra; nedostatečná vazokonstrikce.
11. V jakém věku začíná dítě pociťovat denní teplotní výkyvy, jak se liší od těch u dospělých a v jakém věku dosahují normy pro dospělé?
Na konci 1 měsíce života; jsou nevýznamné a do pěti let dosáhnou normy pro dospělé.
12. Jaká je „komfortní zóna“ teploty dítěte, jaká je teplota uvnitř, jaký je tento ukazatel pro dospělé?
Vnější teplota, při které jsou jednotlivé výkyvy teploty pokožky dítěte nejméně výrazné, se pohybuje v rozmezí 21 – 22 oC, u dospělého 18 – 20 oC.
13. Které termoregulační mechanismy jsou v době narození nejvíce připraveny fungovat? Za jakých podmínek lze u novorozenců aktivovat mechanismy třesové termogeneze?
Zvýšená tvorba tepla, převážně netřesavého původu (vysoký metabolismus), pocení. V podmínkách extrémního mrazu.
14. V jakém poměru by měly být bílkoviny, tuky a sacharidy obsaženy ve stravě dětí ve věku 3 a 6 měsíců, 1 roku, nad 1 rok a dospělých?
Do 3 měsíců – 1: 3: 6; v 6 měsících – 1:2:4. Ve věku od 1 roku – 1:1, 2:4, 6, tedy stejně jako u dospělých.
15. Vyjmenujte znaky metabolismu minerálních solí u dětí. S čím to souvisí?
Dochází k zadržování solí v těle, především ke zvýšené potřebě vápníku, fosforu a železa, což souvisí s růstem organismu.
11 Výměna energie
Nezbytnou podmínkou pro zachování života je, aby organismy přijímaly energii z vnějšího prostředí, a přestože primárním zdrojem energie pro všechno živé je Slunce, jeho záření jsou schopny přímo využívat pouze rostliny. Prostřednictvím fotosyntézy přeměňují energii slunečního záření na energii chemických vazeb. Zvířata a lidé získávají potřebnou energii konzumací rostlinné potravy. (Pro masožravce a částečně i pro všežravce slouží jako zdroj energie jiná zvířata - býložravci.)
Živočichové mohou také přímo přijímat energii ze slunečních paprsků, například poikilotermní živočichové si takto udržují tělesnou teplotu; Teplo (přijímané z vnějšího prostředí a generované v těle samotném) však nelze přeměnit na žádný jiný druh energie. Živé organismy toho na rozdíl od technických zařízení zásadně neumí. Stroj, který využívá energii chemických vazeb (například spalovací motor), ji nejprve přemění na teplo a teprve potom na práci: chemickou energii paliva → teplý → práce (expanze plynu ve válci a pohyb pístu). V živých organismech je možné pouze toto schéma: chemická energie → Práce.
Energie chemických vazeb v molekulách potravních látek je tedy pro živočišný organismus prakticky jediným zdrojem energie a tepelnou energii může využívat pouze k udržení tělesné teploty. Navíc teplo v důsledku rychlého rozptylu v prostředí nemůže být v těle dlouhodobě skladováno. Pokud se v těle objeví přebytečné teplo, stává se to pro homeotermická zvířata vážným problémem a někdy dokonce ohrožuje jejich život (viz část 11.3).
11.1. Zdroje energie a způsoby její přeměny v těle
Živý organismus je otevřený energetický systém: přijímá energii z prostředí (téměř výhradně ve formě chemických vazeb), přeměňuje ji na teplo nebo práci a v této podobě ji vrací zpět do prostředí.
Složky živin, které se dostávají do krve z trávicího traktu (například glukóza, mastné kyseliny nebo aminokyseliny), samy o sobě nejsou schopny přímo přenášet energii svých chemických vazeb na své konzumenty, například drasliko-sodíkovou pumpu nebo sval. aktin a myosin. Mezi potravinovými „nositeli energie“ a „spotřebiteli“ energie existuje univerzální prostředník – adenosintrifosfát (ATP). On je ten pravý přímý zdroj energie pro jakékoli procesy v živých věcech
tělo. Molekula ATP je kombinací adeninu, ribózy a tří fosfátových skupin (obr. 11.1).
Vazby mezi zbytky kyselin (fosfáty) obsahují značné množství energie. Odštěpením koncového fosfátu působením enzymu ATPázy se ATP přemění na adenosindifosfát (ADP). Tím se uvolní 7,3 kcal/mol energie. Energie chemických vazeb v molekulách potravy se využívá k resyntéze ATP z ADP. Uvažujme tento proces s použitím glukózy jako příkladu (obr. 11.2).
První fází využití glukózy je glykolýza Během tohoto procesu se nejprve přemění molekula glukózy na kyselina pyrohroznová (pyruvát), a zároveň poskytuje energii pro resyntézu ATP. Pyruvát se pak převede na acetyl koenzym A - výchozí produkt pro další fázi recyklace - Krebsův cyklus. Mnohonásobné přeměny látek, které tvoří podstatu tohoto cyklu, poskytují další energii pro resyntézu ATP a končí uvolněním vodíkových iontů. Třetí fáze začíná přenosem těchto iontů do dýchacího řetězce - oxidativní fosforylace, v důsledku čehož vzniká i ATP.
Všechny tři stupně recyklace (glykolýza, Krebsův cyklus a oxidativní fosforylace) dohromady tvoří proces tkáňové dýchání. Zásadně důležité je, aby první stupeň (glykolýza) probíhal bez použití kyslíku (anaerobní dýchání) a vede k tvorbě pouze dvou molekul ATP. Dvě následující fáze (Krebsův cyklus a oxidativní fosforylace) mohou probíhat pouze v kyslíkovém prostředí (aerobní dýchání). Kompletní využití jedné molekuly glukózy vede ke vzniku 38 molekul ATP.
Existují organismy, které nejen že nepotřebují kyslík, ale také umírají v kyslíkovém (nebo vzdušném) prostředí - obligátní anaeroby. Patří mezi ně například bakterie způsobující plynovou gangrénu (Clostridium perfringes), tetanus (C. tetani), botulismus (C. botulinum) atd.
U zvířat jsou anaerobní procesy pomocným typem dýchání. Například při intenzivních a častých svalových kontrakcích (nebo při statických kontrakcích) dodávka kyslíku krví zaostává za potřebami svalových buněk. V této době dochází k tvorbě ATP anaerobně s akumulací pyruvátu, který se přeměňuje na kyselina mléčná (laktát). Rostoucí kyslíkový dluh. Zastavením nebo oslabením svalové práce se odstraní rozpor mezi potřebou kyslíku tkáně a možnostmi jeho dodání se laktát přemění na pyruvát, ten se buď přes stadium acetylkoenzymu A oxiduje v Krebsově cyklu na oxid uhličitý, popř. glukoneogenezí se mění na glukózu.
Podle druhého termodynamického zákona dochází k jakékoli přeměně energie z jednoho typu na druhý s obligátním vytvořením značného množství tepla, které se pak rozptýlí v okolním prostoru. K syntéze ATP a přenosu energie z ATP ke skutečným „konzumentům energie“ tedy dochází se ztrátou přibližně poloviny ve formě tepla. Zjednodušeně můžeme tyto procesy znázornit následovně (obr. 11.3).
Přibližně polovina chemické energie obsažené v potravě se okamžitě přemění na teplo a rozptýlí se v prostoru, druhá polovina jde na tvorbu ATP. Při následném rozpadu ATP se polovina uvolněné energie opět přemění na teplo. Výsledkem je, že zvíře a člověk nemohou strávit více než 1/4 veškeré energie spotřebované ve formě potravy na vnější práci (například běh nebo pohyb jakýchkoli předmětů v prostoru). Účinnost vyšších zvířat a lidí (asi 25 %) je tedy několikanásobně vyšší než například účinnost parního stroje.
Veškerá vnitřní práce (kromě procesů růstu a hromadění tuku) se rychle mění v horko. Příklady: a) energie produkovaná srdcem se přeměňuje na teplo v důsledku odporu krevních cév vůči průtoku krve; (b) žaludek vykonává sekreci kyseliny chlorovodíkové, slinivka vylučuje bikarbonátové ionty, v tenkém střevě tyto látky interagují a v nich uložená energie se přeměňuje na teplo.
Výsledky vnější (užitečné) práce vykonávané zvířetem nebo člověkem se také nakonec mění v teplo: pohyb těles v prostoru ohřívá vzduch, vztyčené stavby se hroutí a odevzdávají energii v nich obsaženou zemi a vzduchu ve formě tepla. Egyptské pyramidy jsou vzácným příkladem toho, jak energie svalové kontrakce, vynaložená před téměř 5000 lety, stále čeká na nevyhnutelnou přeměnu v teplo.
Rovnice energetické bilance:
E = A + H + S,
Kde E - celkové množství energie přijaté tělem z potravy; A - externí (užitečná) práce; N - přenos tepla; S- uloženou energii.
Energetické ztráty močí, kožním mazem a dalšími sekrety jsou extrémně malé a lze je zanedbat.
Respirační koeficient je 18,10:24,70 = 0,73.[...]
Dýchací koeficient nezůstává během normálního zrání ovoce konstantní. V premenopauzálním stadiu je to přibližně 1 a dozráváním dosahuje hodnot 1,2... 1,5. Při odchylkách ±0,25 od jedničky ještě nejsou u plodů pozorovány metabolické odchylky a až při velkých odchylkách lze předpokládat fyziologické poruchy. Intenzita dýchání jednotlivých vrstev tkáně žádného plodu není stejná. V souladu s větší aktivitou enzymů v kůži je v ní frekvence dýchání mnohonásobně vyšší než v parenchymatické tkáni (Hulme a Rhodes, 1939). S poklesem obsahu kyslíku a zvýšením koncentrace oxidu uhličitého v parenchymových buňkách klesá intenzita dýchání se vzdáleností od slupky k jádru plodu.[...]
Přístroj na stanovení respiračního koeficientu, pinzeta, proužky filtračního papíru, přesýpací hodiny na 2 minuty, skleněné kelímky, pipety, skleněné tyčinky, 250 ml kónické baňky.[...]
Zařízení pro stanovení respiračního koeficientu se skládá z velké zkumavky s těsně přiléhající pryžovou zátkou, do které se vkládá do pravého úhlu zalomená měřicí trubice se stupnicí milimetrového papíru.[...]
Spotřeba kyslíku a koeficient jeho využití byly konstantní při snížení p02 na 60 a 20 % původní hodnoty (v závislosti na průtoku). Při koncentracích kyslíku mírně nad kritickou úrovní byl maximální objem ventilace udržován po dlouhou dobu (několik hodin). Objem ventilace se zvýšil 5,5krát, ale na rozdíl od kapra se snížil od 22% úrovně nasycení vody kyslíkem. Autoři se domnívají, že snížení objemu ventilace u ryb v extrémní hypoxii je důsledkem kyslíkového deficitu dýchacích svalů. Poměr dechové frekvence a srdeční frekvence byl 1,4 normálně a 4,2 s nedostatkem kyslíku.
Úvodní vysvětlivky. Výhody metody: vysoká citlivost, která umožňuje pracovat s malými vzorky experimentálního materiálu; schopnost pozorovat dynamiku výměny plynů a současně brát v úvahu výměnu plynů 02 a C02, což umožňuje stanovit respirační koeficient.[...]
Proto hodnota pH v oxyteiku klesá na téměř 6,0, zatímco v aerační nádrži pH>7D Při maximální zátěži je příkon pro oxytank včetně výkonu zařízení na výrobu kyslíku 1,3 m3/ (hp-h). a výkonového perlátoru (obr. 26.9), by měl být menší než výkon perlátoru pro provzdušňovací nádrž. To se vysvětluje vysokou koncentrací kyslíku (nad 60 %) ve všech stupních kyslíkové nádrže.[...]
Dynamika uvolňování oxidu uhličitého (С?СО2), absorpce kyslíku ([...]
Mořské a sladkovodní ryby za těchto experimentálních podmínek měly přibližně stejný respirační koeficient (RQ). Nevýhodou těchto údajů je, že autor vzal pro srovnání zlatou rybku, která obecně spotřebovává málo kyslíku a jen stěží může sloužit jako etalon srovnání.[...]
S ohledem na výměnu plynů hibernujícího hmyzu je třeba říci, že klesá i respirační koeficient1. Například Dreyer (1932) zjistil, že v aktivním stavu mravence Formica ulkei Emery byl respirační koeficient 0,874; když byli mravenci před hibernací neaktivní, respirační koeficient se snížil na 0,782 a během období hibernace pokles dosáhl 0,509-0,504. Brambořík z Colorada Leptinotarsa decemlineata Say. v období zimování se respirační koeficient snižuje na 0,492-0,596, zatímco v létě je 0,819-0,822 (Ushatinskaya, 1957). Vysvětluje to skutečnost, že v aktivním stavu se hmyz živí převážně bílkovinami a uhlohydráty, zatímco v hibernaci se spotřebovává hlavně tuk, který vyžaduje méně kyslíku pro oxidaci.
V uzavřených nádobách určených pro tlak v GP RK. d = 1962 Pa (200 mm vodního sloupce), při vysokých rychlostech obměny může být doba nečinnosti nádrže s „mrtvým“ zbytkem před zahájením plnění tak krátká, že se dýchací ventil nestihne otevřít pro „výdech“ “. Pak nedochází ke ztrátám „zpětným výdechem“.[...]
Pro pochopení biochemických procesů probíhajících v těle má velký význam hodnota respiračního koeficientu. Respirační koeficient (RC) je poměr vydechované kyseliny uhličité ke spotřebovanému kyslíku.[...]
Aby bylo možné posoudit vliv teploty na jakýkoli proces, obvykle pracují na hodnotě teplotního koeficientu. Teplotní koeficient (t>ω) procesu dýchání závisí na typu rostliny a na teplotních gradacích. Se zvýšením teploty z 5 na 15 °C se tedy 0 ω může zvýšit na 3, zatímco zvýšení teploty z 30 na 40 °C zvyšuje intenzitu dýchání méně výrazně (ω asi 1,5). Fáze vývoje rostliny je velmi důležitá. Podle B., A. Rubina jsou v každé fázi vývoje rostliny pro proces dýchání nejpříznivější teploty, na jejichž pozadí tato fáze obvykle probíhá Změna optimálních teplot při dýchání rostliny v závislosti na fázi jejich vývoj je dán tím, že v procesu ontogeneze mění dýchací výměnné dráhy. Mezitím jsou různé teploty nejpříznivější pro různé enzymové systémy. V tomto ohledu je zajímavé, že v pozdějších fázích vývoje rostlin jsou pozorovány případy, kdy flavindehydrogenázy působí jako finální oxidázy převádějící vodík přímo na vzdušný kyslík.[...]
Všechny studované ryby v zajetí spotřebují méně kyslíku než v přirozených podmínkách. Mírné zvýšení respiračního koeficientu u ryb chovaných v akváriích ukazuje na změnu kvalitativní stránky metabolismu směrem k větší účasti sacharidů a bílkovin v něm. Autor to vysvětluje horším kyslíkovým režimem akvária oproti přírodním podmínkám; Ryby v akváriu jsou navíc neaktivní.[...]
Pro snížení emisí škodlivých par se také používají reflektorové kotouče, instalované pod montážní trubku dýchacího ventilu. Při vysoké rychlosti obměny atmosférických nádrží může účinnost reflektorových kotoučů dosahovat 20-30%.[...]
K opětovnému nasycení plynové komory může dojít po naplnění, pokud plynový prostor nebyl zcela nasycen parami. V tomto případě se dýchací ventil po naplnění nádoby neuzavře a okamžitě začne další výdech. K tomuto jevu dochází u nádrží, které mají vysoký obratový poměr nebo jsou částečně naplněné, ne do maximální výšky plnění, a dále u nádrží s pomalými procesy sycení hydraulické kapaliny (nádrže s pontony a zapuštěné). Sytost GP je typická zejména pro nádrže, které se plní poprvé po vyčištění a odvětrání. Tento typ ztráty se někdy nazývá ztráty ze saturace nebo saturace GP.[...]
Pro známé u0 Acjcs lze také určit z grafů podobných těm na Obr. 14. Metody pro výpočet ztrát poskytují podobné grafy pro typické nádrže RVS, různé typy dýchacích ventilů a jejich množství. Hodnota Ac/cs znamená nárůst koncentrace v čerpací stanici za celkovou dobu odstávky (tp) a plnění zásobníku (te), tj. t = t„ + t3; určuje se přibližně z grafů (viz obr. 3). Při použití vzorce (!9) je nutné mít na paměti, že při plném nasycení GP ccp/cs = 1 a že doba úplného nasycení GP pozemních nádrží je omezena na 2-4 dny ( v závislosti na povětrnostních podmínkách a dalších podmínkách) a graf je " obr. 3 přibližný. Po získání hodnot ccp/cs>l ze vzorce (19), což znamená počátek úplného nasycení dodávky plynu před konec prostoje nebo konec plnění nádrže, je nutné dosadit ccp/cs = 1.[ ..]
Vyhodnoťme kvantitativní vztahy mezi těmito dvěma proudy plynu. Za prvé, poměr objemu uvolněného oxidu uhličitého k objemu spotřebovaného kyslíku (respirační koeficient) u většiny odpadních vod a aktivovaného kalu je menší než jedna. Za druhé, objemové koeficienty přenosu hmoty pro kyslík a oxid uhličitý jsou blízko sebe. Za třetí, fázová rovnovážná konstanta oxidu uhličitého je téměř 30krát menší než u kyslíku. Za čtvrté, oxid uhličitý není přítomen v kalové směsi pouze v rozpuštěném stavu, ale vstupuje také do chemické interakce s vodou.[...]
Při porovnání obou typů dýchání je markantní nestejný poměr absorpce kyslíku k výdeji oxidu uhličitého. Poměr CO2/O2 se označuje jako respirační koeficient KO.[...]
Pokud při dýchání dojde k oxidaci organických látek s relativně vyšším obsahem kyslíku než u sacharidů, například organické kyseliny - šťavelová, vinná a jejich soli, pak bude respirační koeficient výrazně větší než 1. Bude také větší než 1 v případě kdy část kyslíku, používaného pro mikrobiální dýchání, je odebírána ze sacharidů; nebo při dýchání těch kvasinek, u kterých dochází k alkoholovému kvašení současně s aerobním dýcháním. Pokud spolu s aerobním dýcháním probíhají další procesy, při kterých se používá další kyslík, pak bude respirační koeficient menší než 1. Bude také menší než 1, když látky s relativně nízkým obsahem kyslíku, jako jsou bílkoviny, uhlovodíky atd. ., jsou oxidovány během procesu dýchání Díky tomu, že znáte hodnotu respiračního koeficientu, můžete určit, které látky jsou oxidovány během dýchání.
Nejobecnějším ukazatelem rychlosti oxidace je rychlost dýchání, kterou lze posuzovat podle absorpce kyslíku, uvolňování oxidu uhličitého a oxidace organické hmoty. Další ukazatele respiračního metabolismu: hodnota respiračního koeficientu, poměr glykolytické a pentózofosfátové dráhy štěpení cukrů, aktivita redoxních enzymů. Energetickou účinnost dýchání lze posuzovat podle intenzity oxidativní fosforylace mitochondrií.[...]
Trendy uvedené u jablek Cox Orange týkající se vlivu koncentrací kyslíku a oxidu uhličitého ve vzduchu v komoře platí pro všechny ostatní odrůdy jablek, s výjimkou případů, kdy se dýchací koeficient výrazněji zvyšuje s klesající teplotou.
Hodnota DC závisí na dalších důvodech. V některých tkáních dochází v důsledku obtížného přístupu kyslíku spolu s aerobním dýcháním k anaerobnímu dýchání, které není doprovázeno absorpcí kyslíku, což vede ke zvýšení hodnoty DC. Hodnotu koeficientu určuje také úplnost oxidace dýchacího substrátu. Pokud se v tkáních kromě finálních produktů hromadí i méně oxidované sloučeniny (organické kyseliny), pak DC[...]
Kvantitativní stanovení závislosti výměny plynů u ryb na teplotě bylo provedeno mnoha výzkumníky. Studium této problematiky se ve většině případů omezilo především na kvantitativní stránku dýchání – velikost dechového rytmu, množství spotřeby kyslíku a následně výpočet teplotních koeficientů při různých teplotách.[...]
Pro snížení ztrát odpařováním a znečištěním ovzduší jsou benzinové nádrže vybaveny plynovým potrubím spojujícím vzduchové prostory nádrží, ve kterých jsou skladovány produkty stejné značky, a je instalován společný dýchací ventil. Výše popsané „velké a malé dýchání“, ventilace plynového prostoru, také způsobuje znečištění ovzduší při skladování ropných produktů v zemědělských zařízeních, protože s poměrem obratu tankoviště 4-6 je poměr obratu zásob paliva 10- 20, což znamená pokles v poměru využití nádrží 0,4-0,6. Aby se zabránilo znečištění ovzduší, jsou sklady ropy vybaveny čisticími zařízeními a lapači benzínu a oleje.[...]
Dosud získaná data ukazují, že extrémní teploty způsobují inhibici fyziologického systému, zejména transportu plynů v rybách. Současně se rozvíjí bradykardie, zvyšuje se arytmie, klesá spotřeba kyslíku a rychlost jeho využití. Po těchto změnách ve fungování kardiorespiračního aparátu postupně ustává ventilace žaber a v neposlední řadě přestává fungovat myokard. Anoxie dýchacích svalů a celkový nedostatek kyslíku jsou zřejmě jednou z příčin úhynu ryb v důsledku přehřátí. Zvýšení teploty vede ke zrychlení využití kyslíku a v důsledku toho k poklesu jeho napětí v dorzální aortě, což zase slouží jako signál pro zvýšenou ventilaci žaber.[...]
Před použitím modelu je třeba zkontrolovat jeho kinetické parametry. Validaci modelu čistého kyslíkového systému pro čištění domovních a průmyslových odpadních vod provedli Muller et al. (1) Validace modelu pro čištění domovních odpadních vod používala respirační koeficient R.C 1,0, zatímco pro průmyslové odpadní vody je to 0,85 a dokonce 0,60. Dodatečné ověření chemických interakcí bylo provedeno poměrně nedávno při studiu odpadních vod z celulózky a papírny (obr. 26.6). byl zaznamenán nižší požadavek na růst mikroorganismů, než je tradičně pozorováno v biologických systémech [...].
K vyřešení otázky po podstatě vlivu teploty na metabolismus ryb je nutné znát nejen míru zvýšení či snížení metabolismu se změnou teploty, ale i kvalitativní změny v jednotlivých článcích, které tvoří metabolismu. Kvalitativní stránku metabolismu lze do jisté míry charakterizovat takovými koeficienty jako respirační a amoniakální (poměr uvolněného amoniaku jako konečného produktu metabolismu dusíku ke spotřebovanému kyslíku) (obr. 89).[...]
Z výše uvedené rovnice (4) vyplývá, že poměr konstant pro 02 a CO2 je roven 1,15, t.j. použití techniky měření bilance CO2 by zřejmě umožnilo provádět pozorování při mírně vyšších hodnotách 2. a odpovídajícím způsobem vyšší rychlosti proudění. Tato zdánlivá výhoda však zmizí, pokud předpokládáme, že respirační koeficient je menší než 1. Navíc, jak ukázal Talling 32], přesnost stanovení CO2 v přírodních vodách nemůže být lepší než ± 1 µmol/l (0,044 mg/l), a kyslík - ±0,3 umol/l (0,01 mg/l). V důsledku toho, i když vezmeme respirační koeficient rovný 1, přesnost bilanční metody, založené na zohlednění bilance kyslíku, se ukáže být nejméně třikrát vyšší než při stanovení oxidu uhličitého.[...]
V našich studiích byla s některými doplňky použita morfofyziologická metoda. To umožnilo s dostatečnou přesností (±3,5 %) stanovit množství absorbovaného kyslíku, uvolněného oxidu uhličitého a respirační koeficient (RQ) na celých sazenicích starých 10-12 dní a listech rostlin z polních pokusů. Princip této techniky spočívá v tom, že rostliny umístěné v uzavřené nádobě (speciálně konstruovaná plynová pipeta) s atmosférickým vzduchem mění složení vzduchu v důsledku dýchání. Při znalosti objemu nádoby a stanovení procentuálního složení vzduchu na začátku a na konci experimentu tedy není obtížné vypočítat množství CO2 absorbovaného a uvolněného rostlinami.[...]
Různé rostlinné orgány a pletiva se velmi liší v podmínkách zásobování kyslíkem. V listu proudí kyslík volně téměř do každé buňky. Šťavnaté plody, kořeny, hlízy jsou velmi špatně větrané; jsou špatně propustné pro plyny, nejen pro kyslík, ale i pro oxid uhličitý. Přirozeně se v těchto orgánech proces dýchání přesouvá na anaerobní stranu a dechový koeficient se zvyšuje. V meristematických tkáních je pozorováno zvýšení respiračního koeficientu a posun v procesu dýchání na anaerobní stranu. Různé orgány se tedy vyznačují nejen různou intenzitou, ale i nestejnou kvalitou dechového procesu.[...]
Otázka látek používaných v procesu dýchání je pro fyziology dlouho problémem. I v pracích I.P. Borodina se ukázalo, že intenzita procesu dýchání je přímo úměrná obsahu sacharidů v rostlinných tkáních. To dalo důvod předpokládat, že sacharidy jsou hlavní látkou konzumovanou během dýchání. Při objasňování této problematiky má velký význam stanovení respiračního koeficientu. Respirační koeficient je objemový nebo molární poměr CO2 uvolněného při dýchání k CO2 absorbovanému za stejnou dobu Při normálním přístupu kyslíku závisí hodnota respiračního koeficientu na substrátu dýchání. Pokud jsou v procesu dýchání použity sacharidy, pak proces probíhá podle rovnice CeH) 2O5 + 6O2 = 6CO2 + 6H2O, v tomto případě je respirační koeficient roven jedné = 1. Pokud však více oxidovaných sloučenin, jako jsou organické kyseliny, podléhá rozkladu během dýchání, absorpce kyslíku se snižuje a respirační koeficient je větší než jedna. Když je během dýchání oxidováno více redukovaných sloučenin, jako jsou tuky nebo bílkoviny, je potřeba více kyslíku a respirační koeficient je menší než jedna.[...]
Nejjednodušší proces aerobního dýchání je tedy znázorněn v následující formě. Molekulární kyslík spotřebovaný při dýchání se využívá především k vázání vodíku vznikajícího při oxidaci substrátu. Vodík ze substrátu se přenáší na kyslík prostřednictvím řady mezilehlých reakcí, které probíhají postupně za účasti enzymů a nosičů. Takzvaný respirační koeficient dává určitou představu o povaze procesu dýchání. To je chápáno jako poměr objemu uvolněného oxidu uhličitého k objemu kyslíku absorbovaného během dýchání (C02:02).[...]
Výkonnost kardiorespiračního aparátu ryb, jeho rezervní schopnosti a labilita frekvenčních a amplitudových parametrů závisí na druhu a ekologických vlastnostech ryb. Při zvýšení teploty o stejnou hodnotu (z 5 na 20°C) se zvýšila dechová frekvence candáta z 25 na 50 za minutu, u štik ze 46 na 75 au ide z 63 na 112 za minutu. Spotřeba kyslíku se zvyšuje souběžně se zvyšující se frekvencí, ale ne hloubkou dýchání. Největší počet dechových pohybů k čerpání jednotkového objemu vody produkuje mobilní ide a nejméně méně aktivní oxyfilní candát, což pozitivně koreluje s intenzitou výměny plynů u studovaného druhu. Poměr maximálního objemu ventilace a odpovídajícího koeficientu využití kyslíku podle autorů určuje maximální energetické možnosti organismu. V klidu byla nejvyšší intenzita výměny plynů a objem ventilace u candátů oxyfilních a při funkční zátěži (motorická aktivita, hypoxie) - v ide. Při nízkých teplotách bylo zvýšení ventilačního objemu in ide v reakci na hypoxii větší než při vysokých teplotách, konkrétně: 20krát při 5 °C a 8krát při 20 °C. U Orthologus thioglossy se při hypoxii (40% nasycení) objem vody čerpané žábrami mění v menší míře: při 12°C se zvětší 5x a při 28°C 4,3x.[...]
Ukazatele metabolismu sacharidů při adaptivní exogenní hypoxii, tj. při mírném až středním nedostatku kyslíku v prostředí, byly prostudovány mnohem méně. Omezená dostupná experimentální data však ukazují, že v tomto případě dochází ke zvýšenému využití glykogenu ve svalech, zvýšení kyseliny mléčné a cukru v krvi. Jak by se dalo očekávat, úroveň nasycení vody kyslíkem, při které jsou tyto posuny pozorovány, není u různých druhů stejná. Například u mihule byla pozorována hyperglykémie, když se obsah kyslíku snížil pouze o 20 % původní úrovně a u 1 abea karepvka zůstala koncentrace krevního cukru trvale nízká i při 40% nasycení vody kyslíkem a pouze další snížení saturace vedlo k rychlému zvýšení hladiny cukru v krvi. Při hypoxii u lína bylo zaznamenáno zvýšení hladiny cukru v krvi a kyseliny mléčné. Podobná reakce na hypoxii byla zaznamenána u kanálových sumců. V první z těchto studií bylo při 50% nasycení vody kyslíkem zjištěno u ryb zvýšení obsahu kyseliny mléčné, které pokračovalo v první hodině normoxie, tedy po návratu ryb do normálních kyslíkových podmínek. K obnovení biochemických parametrů do normálu došlo během 2-6 hodin a zvýšení obsahu laktátu a respiračního koeficientu z 0,8 na 2,0 indikovalo zvýšení anaerobní glykolýzy.
Respirační kvocient (RC) je poměr objemu uvolněného oxidu uhličitého k objemu kyslíku absorbovaného za určitou dobu. Pokud se během metabolického procesu v těle oxidují pouze sacharidy, pak bude respirační koeficient roven 1. To lze vidět z následujícího vzorce:
V důsledku toho je k vytvoření jedné molekuly CO2 během metabolismu sacharidů zapotřebí jedna molekula O2. Protože podle Avogadro-Gerardova zákona zaujímá stejný počet molekul při stejné teplotě a tlaku stejné objemy. Proto bude respirační koeficient pro oxidaci sacharidů roven 1:
![]()
Pro tuky to bude:
![]()
Oxidace jedné molekuly tuku vyžaduje 81,5 molekul kyslíku a oxidace 1 gramu molekuly tuku vyžaduje 81,5 x 22,4 litrů kyslíku, tedy 1825,6 litrů O 2, kde 22,4 je objem jednoho gramu molekuly v litrech. . Gram molekuly tuku se rovná 890 g, pak oxiduje 1 litr kyslíku ![]() 487 g tuku. 1 g tuku po úplné oxidaci uvolní 38,945 kJ (9,3 kcal)* a 0,487 dává 18,551 kJ. Kalorický ekvivalent 1 litru kyslíku s respiračním koeficientem 0,7 se tedy bude rovnat 18,551 kJ. Za normálních podmínek leží respirační koeficient mezi 1 a 0,7. Při DC 0,7 se v těle oxidují tuky a kalorický ekvivalent neboli kalorická hodnota 1 litru kyslíku je 18,551 kJ a při DC 1 je 21,135.
487 g tuku. 1 g tuku po úplné oxidaci uvolní 38,945 kJ (9,3 kcal)* a 0,487 dává 18,551 kJ. Kalorický ekvivalent 1 litru kyslíku s respiračním koeficientem 0,7 se tedy bude rovnat 18,551 kJ. Za normálních podmínek leží respirační koeficient mezi 1 a 0,7. Při DC 0,7 se v těle oxidují tuky a kalorický ekvivalent neboli kalorická hodnota 1 litru kyslíku je 18,551 kJ a při DC 1 je 21,135.