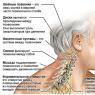
Stanovenie respiračného koeficientu (RK) rastlín. Stanovenie respiračného koeficientu Respiračný koeficient jeho hodnota
respiračný koeficient (RK)
pomer objemu oxidu uhličitého uvoľneného cez pľúca k objemu kyslíka absorbovaného za rovnaký čas; hodnota D.c., keď je subjekt v pokoji, závisí od druhu potravín oxidovaných v tele.
Encyklopedický slovník, 1998
respiračný kvocient
pomer objemu oxidu uhličitého uvoľneného pri dýchaní za určitý čas k objemu kyslíka absorbovaného za rovnaký čas. Charakterizuje vlastnosti výmeny plynov a metabolizmu u zvierat a rastlín. U zdravého človeka je to približne 0,85.
Respiračný koeficient
pomer objemu oxidu uhličitého uvoľneného z tela k objemu kyslíka absorbovaného za rovnaký čas. Označené:
Stanovenie DC je dôležité pre štúdium charakteristík výmeny plynov a metabolizmu u zvierat a rastlinných organizmov. Keď sú sacharidy v tele oxidované a kyslík je plne dostupný, DC je 1, tuky ≈ 0,7, bielkoviny ≈ 0,8. U zdravého človeka v pokoji je DC 0,85 ╠ 0,1; pri miernej práci, ako aj u zvierat, ktoré sa živia prevažne rastlinnou potravou, sa približuje k 1. U ľudí pri veľmi dlhej práci, pôste, u mäsožravcov (predátorov), ako aj pri zimnom spánku, kedy v dôsledku obmedzených zásob sacharidov v tele, disimilácia zvyšuje tuk, DC je asi 0,7. DC presahuje 1 pri intenzívnom ukladaní tukov v tele zo sacharidov dodávaných s potravou (napr. u ľudí pri obnove normálnej hmotnosti po pôste, po dlhodobých chorobách, ako aj u zvierat pri výkrme). DC sa zvyšuje na 2 pri zvýšenej práci a hyperventilácii pľúc, kedy sa z tela uvoľňuje ďalší CO2, ktorý bol vo viazanom stave. Ešte väčšie hodnoty DC dosahuje v anaeróboch, v ktorých väčšina uvoľneného CO2 vzniká oxidáciou bez kyslíka (fermentáciou). DK pod 0,7 sa vyskytuje pri ochoreniach spojených s poruchami metabolizmu, po ťažkej fyzickej práci.
U rastlín závisí DK od chemickej povahy dýchacieho substrátu, obsahu CO2 a O2 v atmosfére a ďalších faktorov, čím charakterizuje špecifiká a podmienky dýchania. Keď bunka používa sacharidy na dýchanie (semenáčiky zrna), DC je približne 1, tuky a bielkoviny (klíčiace olejnaté semená a strukoviny) ≈ 0,4≈0,7. Pri nedostatku O2 a ťažkom prístupe (semená s tvrdou škrupinou) je DC 2≈3 alebo viac; vysoký DC je tiež charakteristický pre bunky rastového bodu.
(DC) je pomer objemu oxidu uhličitého uvoľneného počas dýchania k objemu absorbovaného kyslíka.
Hodnota koeficientu dýchania rastlín
hodnota DC označuje povahu materiálu oxidovaného počas dýchania a typ dýchania; môže byť rovná, väčšia alebo menšia ako jedna. Počas oxidácie uhľohydrátov sú objemy vymenených plynov, oxidu uhličitého a kyslíka, rovnaké a pomer C0 2: 0 2 sa rovná jednotke. V tomto prípade sa kyslík spotrebovaný pri dýchaní využíva len na oxidáciu uhlíka na oxid uhličitý, pretože pomer vodíka a kyslíka v molekule glukózy je taký, že na oxidáciu vodíka na vodu je v cukre dostatok kyslíka. samotná molekula. Keď sa oxiduje množstvo organických kyselín, respiračný koeficient rastlín je väčší ako jedna. Kyselina šťaveľová je teda zlúčenina bohatšia na kyslík ako sacharidy. Kyslík prítomný v molekule nestačí len na oxidáciu vodíka na vodu, ale časť z neho zostáva na oxidáciu uhlíka; preto na úplnú oxidáciu dvoch molekúl kyseliny šťaveľovej stačí jedna molekula kyslíka: 2C 2 H 2 O 4 + O 2 → 4CO 2 + 2H 2 O, DC (4CO 2: O 2) sa v tomto prípade rovná až 4. V prípadoch, keď rastlina dýcha vďaka bielkovinám alebo tukom, ktorých molekula obsahuje veľa vodíka a uhlíka a málo kyslíka DC je menej ako jedna, keďže na oxidáciu všetkého uhlíka a vodíka, ktoré sa nachádzajú v týchto zlúčeninách; , je potrebné absorbovať veľké množstvo kyslíka. Keď sa kyselina stearová oxiduje, oxidačná reakcia bude prebiehať nasledovne: C18H2602 + 2602 → 18C02 + 18H20. DC (18C02:2602) je 0,69. V prípade oxidácie sacharidov sa teda DC rovná jednej, organické kyseliny - viac ako jedna, bielkoviny a tuky - menej ako jedna.
Tepelný účinok počas dýchania rastlín
Tepelný efekt bude mať opačnú hodnotu DC: maximálny tepelný efekt bude pri oxidácii tukov, pretože ide o najviac redukované zlúčeniny. Závislosť hodnoty DC od charakteru dýchacieho materiálu sa pozoruje len vtedy, keď je v prostredí a rastlinných tkanivách dostatok kyslíka. Avšak počas oxidácie toho istého dýchacieho materiálu, ale s nedostatkom kyslíka v prostredí a rastlinných tkanivách, sa môžu zmeniť aj hodnoty DC. Ak je kyslíka málo, tak oxidácia neprebehne úplne a okrem oxidu uhličitého a vody vznikajú organické kyseliny, ktoré sú viac oxidované ako sacharidy. V tomto prípade bude DC menšia ako jedna, pretože časť absorbovaného kyslíka zostane v molekulách vytvorených organických kyselín, zatiaľ čo sa uvoľní menej oxidu uhličitého. Uvoľní sa menej energie, pretože časť z nej bude uložená v organických kyselinách.1. Aký proces zabezpečuje uvoľňovanie energie v tele? Čo je jej podstatou?
Disimilácia (katabolizmus), t.j. rozpad bunkových štruktúr a zlúčenín tela s uvoľňovaním energie a produktov rozpadu.
2. Aké živiny dodávajú telu energiu?
Sacharidy, tuky a bielkoviny.
3. Vymenujte hlavné metódy stanovenia množstva energie vo vzorke výrobku.
Fyzikálna kalorimetria; fyzikálno-chemické metódy na stanovenie množstva živín vo vzorke s následným výpočtom energie v nej obsiahnutej; podľa tabuliek.
4. Popíšte podstatu metódy fyzikálnej kalorimetrie.
Vzorka produktu sa spáli v kalorimetri a následne sa vypočíta uvoľnená energia na základe stupňa ohrevu vody a materiálu kalorimetra.
5. Napíšte vzorec na výpočet množstva tepla uvoľneného pri spaľovaní produktu v kalorimetri. Dešifrujte jeho symboly.
Q = MvSv (t2 - t1) + MkSk (t2 - t1) - Qo,
kde Q je množstvo tepla, M je hmotnosť (w - voda, k - kalorimeter), (t 2 - t 1) je teplotný rozdiel medzi vodou a kalorimetrom po a pred spálením vzorky, C je špecifické teplo kapacita, Qo je množstvo tepla generovaného oxidačným činidlom.
6. Aké sú fyzikálne a fyziologické kalorické koeficienty živiny?
Množstvo tepla uvoľneného pri spaľovaní 1 g látky v kalorimetri a v tele, resp.
7. Koľko tepla sa uvoľní, keď sa v kalorimetri spáli 1 g bielkovín, tukov a sacharidov?
1g bielkovín – 5,85 kcal (24,6 kJ), 1g tuku – 9,3 kcal (38,9 kJ), 1g sacharidov – 4,1 kcal (17,2 kJ).
8. Formulujte Hessov termodynamický zákon, na základe ktorého sa vypočíta energia vstupujúca do tela na základe množstva strávených bielkovín, tukov a sacharidov.
Termodynamický efekt závisí len od obsahu tepla v počiatočných a konečných reakčných produktoch a nezávisí od intermediárnych premien týchto látok.
9. Koľko tepla sa uvoľní pri oxidácii 1 g bielkovín, 1 g tukov a 1 g sacharidov v tele?
1 g bielkovín – 4,1 kcal (17,2 kJ), 1 g tukov – 9,3 kcal (38,9 kJ), 1 g sacharidov – 4,1 kcal (17,2 kJ).
10. Vysvetlite príčinu rozdielu medzi fyzikálnym a fyziologickým kalorickým koeficientom pre bielkoviny. V ktorom prípade je to väčšie?
V kalorimetri (fyzikálny koeficient) sa bielkovina rozkladá na konečné produkty - CO 2, H 2 O a NH 3 za uvoľnenia všetkej energie v nich obsiahnutej. V tele (fyziologický koeficient) sa bielkoviny rozkladajú na CO 2, H 2 O, močovinu a ďalšie látky metabolizmu bielkovín, ktoré obsahujú energiu a vylučujú sa močom.
Stanoví sa obsah bielkovín, tukov a uhľohydrátov v potravinách, ich množstvo sa vynásobí príslušnými fyziologickými kalorickými koeficientmi, sčíta sa a od súčtu sa odpočíta 10 %, ktoré sa nevstrebávajú v tráviacom trakte (straty stolicou).
12. Vypočítajte (v kcal a kJ) energetický príjem pri príjme 10 g bielkovín, tukov a sacharidov do tela spolu s jedlom.
Q = 4,110 + 9,310 + 4,110 = 175 kcal. (175 kcal - 17,5 kcal) x 4,2 kJ, kde 17,5 kcal je energia nestrávených živín (straty stolicou - cca 10%). Spolu: 157,5 kcal (661,5 kJ).
Kalorimetria: priama (metóda Atwater-Benedict); nepriame, alebo nepriame (metódy Krogh, Shaternikov, Douglas - Holden).
14. Na čom je založený princíp priamej kalorimetrie?
Pri priamom meraní množstva tepla generovaného telom.
15. Stručne opíšte konštrukciu a princíp fungovania kamery Atwater-Benedict.
Komora, v ktorej je testovaný subjekt, je tepelne izolovaná od okolia, jej steny neabsorbujú teplo, sú to radiátory, cez ktoré prúdi voda. Na základe stupňa ohrevu určitého množstva vody sa vypočíta množstvo tepla spotrebovaného telom.
16. Na čom je založený princíp nepriamej (nepriamej) kalorimetrie?
Výpočtom množstva uvoľnenej energie podľa údajov o výmene plynu (absorbovaný O 2 a uvoľnený CO 2 za deň).
17. Prečo možno množstvo energie uvoľnenej telom vypočítať na základe výmenných kurzov plynov?
Pretože množstvo telom spotrebovaného O 2 a uvoľneného CO 2 presne zodpovedá množstvu zoxidovaných bielkovín, tukov a sacharidov, a teda aj telu spotrebovanej energii.
18. Aké koeficienty sa používajú na výpočet spotreby energie nepriamou kalorimetriou?
Respiračný koeficient a kalorický ekvivalent kyslíka.
19. Čo sa nazýva respiračný koeficient?
Pomer objemu oxidu uhličitého uvoľneného telom k objemu kyslíka spotrebovaného za rovnaký čas.
20. Vypočítajte respiračný koeficient (RC), ak je známe, že vdychovaný vzduch obsahuje 17 % kyslíka a 4 % oxidu uhličitého.
Keďže atmosférický vzduch obsahuje 21 % O 2 , percento absorbovaného kyslíka je 21 % - 17 %, t.j. 4 %. CO 2 vo vydychovanom vzduchu je tiež 4 %. Odtiaľto
21. Od čoho závisí dýchací koeficient?
22. Aký je respiračný koeficient pri oxidácii v organizme na konečné produkty bielkovín, tukov a sacharidov?
Počas oxidácie bielkovín – 0,8, tukov – 0,7, sacharidov – 1,0.
23. Prečo je respiračný kvocient nižší pre tuky a bielkoviny ako pre sacharidy?
Viac O2 sa spotrebuje na oxidáciu bielkovín a tukov, pretože obsahujú menej intramolekulárneho kyslíka ako uhľohydráty.
24. Akú hodnotu má dýchací kvocient človeka na začiatku intenzívnej fyzickej práce? prečo?
Do jedného, pretože zdrojom energie sú v tomto prípade najmä sacharidy.
25. Prečo je dýchací koeficient človeka väčší ako jeden v prvých minútach po intenzívnej a dlhšej fyzickej práci?
Pretože sa uvoľní viac CO 2 ako sa spotrebuje O 2, pretože kyselina mliečna nahromadená vo svaloch vstupuje do krvi a vytláča CO 2 z bikarbonátov.
26. Čo sa nazýva kalorický ekvivalent kyslíka?
Množstvo tepla uvoľneného telom pri konzumácii 1 litra O2.
27. Od čoho závisí kalorický ekvivalent kyslíka?
Z pomeru bielkovín, tukov a sacharidov zoxidovaných v tele.
28. Aký je kalorický ekvivalent kyslíka pri oxidácii v tele (v procese disimilácie) bielkovín, tukov a sacharidov?
Na bielkoviny – 4,48 kcal (18,8 kJ), na tuky – 4,69 kcal (19,6 kJ), na sacharidy – 5,05 kcal (21,1 kJ).
29. Stručne opíšte proces stanovenia spotreby energie metódou Douglas-Holden (úplná analýza plynu).
V priebehu niekoľkých minút subjekt vdýchne atmosférický vzduch a vydýchnutý vzduch sa zhromažďuje do špeciálneho vaku, meria sa jeho množstvo a vykonáva sa analýza plynov, aby sa určil objem spotrebovaného kyslíka a uvoľneného CO 2 . Vypočíta sa respiračný koeficient, pomocou ktorého sa z tabuľky zistí zodpovedajúci kalorický ekvivalent O 2, ktorý sa následne vynásobí objemom O 2 spotrebovaným za dané časové obdobie.
30. Stručne opíšte metódu M. N. Shaternikova na stanovenie energetického výdaja u zvierat v experimente.
Zviera sa umiestni do komory, do ktorej sa dodáva kyslík, keď sa spotrebuje. CO 2 uvoľnený pri dýchaní je absorbovaný alkáliami. Uvoľnená energia sa vypočíta na základe množstva spotrebovaného O2 a priemerného kalorického ekvivalentu O2: 4,9 kcal (20,6 kJ).
31. Vypočítajte spotrebu energie za 1 minútu, ak je známe, že subjekt spotreboval 300 ml O 2 . Dýchací koeficient je 1,0.
DK = 1,0, zodpovedá kalorickému ekvivalentu kyslíka 5,05 kcal (21,12 kJ). Preto spotreba energie za minútu = 5,05 kcal x 0,3 = 1,5 kcal (6,3 kJ).
32. Stručne opíšte proces stanovenia spotreby energie pomocou Kroghovej metódy u ľudí (neúplná analýza plynov).
Subjekt inhaluje kyslík z metabolimetrického vaku, vydychovaný vzduch sa vracia do toho istého vaku, ktorý predtým prešiel cez absorbér CO2. Na základe údajov z metabolimetra sa určí spotreba O2 a vynásobí sa kalorickým ekvivalentom kyslíka 4,86 kcal (20,36 kJ).
33. Vymenujte hlavné rozdiely vo výpočte spotreby energie pomocou metódy Douglas-Holden a Krogh.
Douglas-Holdenova metóda zahŕňa výpočet spotreby energie na základe údajov z kompletnej analýzy plynu; Kroghova metóda - len objemom spotrebovaného kyslíka pomocou kalorického ekvivalentu kyslíka charakteristického pre bazálne metabolické podmienky.
34. Čo sa nazýva bazálny metabolizmus?
Minimálna spotreba energie, ktorá zaisťuje homeostázu za štandardných podmienok: v bdelom stave, s maximálnym svalovým a emocionálnym odpočinkom, nalačno (12 - 16 hodín bez jedla), pri pohodlnej teplote (18 - 20 C).
35. Prečo sa bazálny metabolizmus určuje za štandardných podmienok: maximálny svalový a emocionálny odpočinok, nalačno, pri pohodlnej teplote?
Pretože fyzická aktivita, emočný stres, príjem potravy a zmeny teploty okolia zvyšujú intenzitu metabolických procesov v organizme (spotrebu energie).
36. Aké procesy spotrebúvajú energiu bazálneho metabolizmu v organizme?
Na zabezpečenie životných funkcií všetkých orgánov a tkanív tela, bunkovú syntézu a udržiavanie telesnej teploty.
37. Aké faktory určujú hodnotu správneho (priemerného) bazálneho metabolizmu zdravého človeka?
Pohlavie, vek, výška a telesná hmotnosť (hmotnosť).
38. Aké faktory okrem pohlavia, hmotnosti, výšky a veku určujú hodnotu skutočného (skutočného) bazálneho metabolizmu zdravého človeka?
Životné podmienky, ktorým je telo prispôsobené: trvalý pobyt v chladnom klimatickom pásme zvyšuje bazálny metabolizmus; dlhodobá vegetariánska strava – redukuje.
39. Uveďte spôsoby, ako určiť množstvo správneho bazálneho metabolizmu u človeka. Aká metóda sa používa na určenie hodnoty skutočného bazálneho metabolizmu človeka v praktickej medicíne?
Podľa tabuliek, podľa vzorcov, podľa nomogramov. Kroghova metóda (neúplná analýza plynov).
40. Akú hodnotu má bazálny metabolizmus u mužov a žien za deň, ako aj na 1 kg telesnej hmotnosti za deň?
U mužov 1500 – 1700 kcal (6300 – 7140 kJ), alebo 21 – 24 kcal (88 – 101 kJ)/kg/deň. Ženy majú približne o 10 % menej ako táto hodnota.
41. Je bazálny metabolizmus prepočítaný na 1 m 2 povrchu tela a na 1 kg telesnej hmotnosti rovnaký u teplokrvníkov a ľudí?
Pri prepočte na 1 m 2 povrchu tela u teplokrvných živočíchov rôznych druhov a ľudí sú ukazovatele približne rovnaké, pri prepočte na 1 kg hmotnosti sú veľmi odlišné.
42. Čo sa nazýva pracovná burza?
Kombinácia bazálneho metabolizmu a dodatočného energetického výdaja, ktorý zabezpečuje fungovanie organizmu v rôznych podmienkach.
43. Uveďte faktory, ktoré zvyšujú spotrebu energie v tele. Čo sa nazýva špecifický dynamický efekt jedla?
Fyzický a psychický stres, emočný stres, zmeny teplôt a iných podmienok prostredia, špecifické dynamické účinky potravy (zvýšená spotreba energie po jedle).
44. O koľko percent sa zvýši spotreba energie v tele po konzumácii bielkovín a zmiešaných potravín, tukov a sacharidov?
Po konzumácii bielkovinových potravín - o 20 - 30%, zmiešaných potravín - o 10 - 12%.
45. Ako ovplyvňuje teplota okolia energetický výdaj organizmu?
Zmeny teplôt v rozmedzí 15 – 30 °C výrazne neovplyvňujú energetickú spotrebu organizmu. Pri teplotách pod 15C a nad 30C sa spotreba energie zvyšuje.
46. Ako sa mení metabolizmus pri okolitých teplotách pod 15? čo na tom záleží?
Zvyšovanie. Tým sa zabráni ochladzovaniu tela.
47. Ako sa nazýva výkonnosť tela pri svalovej práci?
Vyjadrený v percentách, pomer energie ekvivalentnej užitočnej mechanickej práci k celkovej energii vynaloženej na vykonanie tejto práce.
48. Uveďte vzorec na výpočet koeficientu výkonnosti (výkonnosti) u človeka pri svalovej práci, uveďte jeho priemernú hodnotu, dešifrujte prvky vzorca.

kde A je energia ekvivalentná užitočnej práci, C je celková spotreba energie, e je spotreba energie za rovnaký čas v pokoji. Účinnosť je 20%.
49. Ktoré živočíchy sa nazývajú poikilotermné a homeotermné?Poikilotermné živočíchy (studenokrvné) - s nestabilnou telesnou teplotou v závislosti od teploty okolia; homeotermické (teplokrvné) - živočíchy s konštantnou telesnou teplotou, ktorá nezávisí od teploty okolia.
50. Aký význam má pre organizmus stálosť telesnej teploty? V ktorých orgánoch prebieha proces tvorby tepla najintenzívnejšie?
Poskytuje vysokú úroveň životnej aktivity relatívne bez ohľadu na okolitú teplotu. Vo svaloch, pľúcach, pečeni, obličkách.
51. Vymenujte druhy termoregulácie. Formulujte podstatu každého z nich.
Chemická termoregulácia – regulácia telesnej teploty zmenou intenzity tvorby tepla; fyzikálna termoregulácia – zmenou intenzity prenosu tepla.
52. Aké procesy zabezpečujú prenos tepla?
Sálanie tepla (sálanie), odparovanie tepla, vedenie tepla, prúdenie.
53. Ako sa mení lúmen krvných ciev kože, keď sa teplota okolia znižuje a zvyšuje? Aký je biologický význam tohto javu?
Keď teplota klesne, cievy v koži sa zúžia. Keď teplota okolia stúpa, cievy v koži sa rozširujú. Faktom je, že zmena šírky lúmenu krvných ciev, regulácia prenosu tepla, pomáha udržiavať konštantnú telesnú teplotu.
54. Ako a prečo sa pri silnej stimulácii sympatoadrenálneho systému mení tvorba a prenos tepla?
Produkcia tepla sa zvýši stimuláciou oxidačných procesov a zníži sa prenos tepla v dôsledku zúženia kožných ciev.
55. Uveďte oblasti lokalizácie termoreceptorov.
Koža, kožné a podkožné cievy, vnútorné orgány, centrálny nervový systém.
56. V ktorých častiach a štruktúrach centrálneho nervového systému sa nachádzajú termoreceptory?
V hypotalame retikulárna formácia stredného mozgu, v mieche.
57. V ktorých častiach centrálneho nervového systému sa nachádzajú centrá termoregulácie? Ktorá štruktúra centrálneho nervového systému je najvyšším centrom termoregulácie?
V hypotalame a mieche. Hypotalamus.
58. Aké zmeny nastanú v organizme pri dlhodobom nedostatku tukov a sacharidov v strave, ale pri optimálnom príjme bielkovín z potravy (80 - 100 g denne)? prečo?
Spotreba dusíka v organizme bude prevýšená nad príjmom a chudnutím, keďže náklady na energiu budú kryté najmä bielkovinami a tukovými zásobami, ktoré sa nedoplňujú.
59. V akom množstve a pomere by mali byť bielkoviny, tuky a sacharidy obsiahnuté v strave dospelého človeka (priemerná verzia)?
Bielkoviny – 90 g, tuky – 110 g, sacharidy – 410 g, pomer 1:1, 2:4, 6.
60. Ako sa mení stav tela s nadmerným príjmom tukov?
Rozvíja sa (predčasne) obezita a ateroskleróza. Obezita je rizikovým faktorom pre rozvoj srdcovo-cievnych ochorení a ich komplikácií (infarkt myokardu, cievna mozgová príhoda a pod.) a zníženia strednej dĺžky života.
1. Aký je pomer hodnôt bazálneho metabolizmu u detí prvých 3–4 rokov života, v období puberty, vo veku 18–20 rokov a dospelých (kcal/kg/deň)?
Do 3–4 rokov majú deti približne 2-krát viac, v puberte – 1,5-krát viac ako dospelí. Vo veku 18–20 rokov zodpovedá norme pre dospelých.
2. Nakreslite graf zmien bazálneho metabolizmu u chlapcov s vekom (u dievčat je bazálny metabolizmus o 5 % nižší).

3. Čo vysvetľuje vysokú intenzitu oxidačných procesov u dieťaťa?
Vyššia úroveň metabolizmu mladých tkanív, relatívne veľký povrch tela a samozrejme väčší výdaj energie na udržanie konštantnej telesnej teploty, zvýšená sekrécia hormónov štítnej žľazy a norepinefrínu.
4. Ako sa menia náklady na energiu na rast v závislosti od veku dieťaťa: do 3 mesiacov života, pred nástupom puberty, počas puberty?
V prvých 3 mesiacoch po narodení sa zväčšujú, potom postupne klesajú a počas puberty sa opäť zvyšujú.
5. Z čoho pozostáva celkový energetický výdaj 1-ročného dieťaťa a ako je percentuálne rozdelený v porovnaní s dospelým?
U dieťaťa: 70 % pripadá na bazálny metabolizmus, 20 % na pohyb a udržiavanie svalového tonusu, 10 % na špecifický dynamický účinok potravy. U dospelého: 50 – 40 – 10 %, resp.
6. Vynakladajú dospelí alebo deti vo veku 3–5 rokov viac energie pri vykonávaní svalovej práce na dosiahnutie rovnakého priaznivého výsledku, koľkokrát a prečo?
Deti 3 až 5-krát, pretože majú menej dokonalú koordináciu, čo vedie k nadmerným pohybom, čo má za následok výrazne menej užitočnú prácu pre deti.
7. Ako sa mení výdaj energie, keď dieťa plače, o koľko percent a v dôsledku čoho?
Zvyšuje sa o 100–200 % v dôsledku zvýšenej produkcie tepla v dôsledku emocionálneho vzrušenia a zvýšenej svalovej aktivity.
8. Akú časť (v percentách) energetického výdaja dojčaťa zabezpečujú bielkoviny, tuky a sacharidy? (porovnaj s normou pre dospelých).
Kvôli bielkovinám - 10%, kvôli tukom - 50%, kvôli sacharidom - 40%. U dospelých – 20 – 30 – 50 %, resp.
9. Prečo sa deti, najmä v dojčenskom veku, rýchlo prehrievajú, keď teplota okolia stúpa? Znášajú deti zvýšenie alebo zníženie teploty okolia ľahšie?
Pretože deti majú zvýšenú tvorbu tepla, nedostatočné potenie a teda aj odparovanie tepla, nezrelé centrum termoregulácie. Degradácia.
10. Pomenujte bezprostrednú príčinu a vysvetlite mechanizmus rýchleho ochladzovania detí (najmä dojčiat) pri poklese okolitej teploty.
Zvýšený prenos tepla u detí v dôsledku pomerne veľkého povrchu tela, bohatého prekrvenia pokožky, nedostatočnej tepelnej izolácie (tenká koža, nedostatok podkožného tuku) a nezrelosti centra termoregulácie; nedostatočná vazokonstrikcia.
11. V akom veku začína dieťa pociťovať denné teplotné výkyvy, ako sa líšia od tých u dospelých a v akom veku dosahujú normy pre dospelých?
Na konci 1 mesiaca života; sú nevýznamné a do piatich rokov dosiahnu normu pre dospelých.
12. Aká je „komfortná zóna“ teploty dieťaťa, aká je v ňom teplota, aký je tento ukazovateľ pre dospelých?
Vonkajšia teplota, pri ktorej sú jednotlivé výkyvy teploty pokožky dieťaťa najmenej výrazné, je v rozmedzí 21 – 22 oC, u dospelého 18 – 20 oC.
13. Ktoré termoregulačné mechanizmy sú najpripravenejšie fungovať v čase narodenia? Za akých podmienok sa u novorodencov môžu aktivovať mechanizmy chvenia termogenézy?
Zvýšená tvorba tepla, prevažne netrasúceho sa pôvodu (vysoký metabolizmus), potenie. V podmienkach extrémneho chladu.
14. V akom pomere by mali byť bielkoviny, tuky a sacharidy obsiahnuté v strave detí vo veku tri a šesť mesiacov, 1 rok, viac ako jeden rok a dospelých?
Do 3 mesiacov – 1: 3: 6; vo veku 6 mesiacov – 1: 2: 4. Vo veku 1 rok a viac – 1: 1, 2: 4, 6, t.j. rovnako ako u dospelých.
15. Vymenujte znaky metabolizmu minerálnych solí u detí. S čím to súvisí?
V organizme dochádza k zadržiavaniu solí, najmä k zvýšenej potrebe vápnika, fosforu a železa, čo súvisí s rastom organizmu.
11 Výmena energie
Nevyhnutnou podmienkou pre udržanie života je, aby organizmy prijímali energiu z vonkajšieho prostredia, a hoci primárnym zdrojom energie pre všetko živé je Slnko, len rastliny sú schopné jeho žiarenie priamo využívať. Prostredníctvom fotosyntézy premieňajú energiu slnečného žiarenia na energiu chemických väzieb. Zvieratá a ľudia získavajú potrebnú energiu konzumáciou rastlinnej potravy. (Pre mäsožravce a čiastočne pre všežravce slúžia ako zdroj energie iné živočíchy - bylinožravce.)
Živočíchy môžu priamo prijímať energiu aj zo slnečných lúčov, napríklad poikilotermné živočíchy si takto udržiavajú telesnú teplotu; Teplo (prijímané z vonkajšieho prostredia a generované v samotnom tele) sa však nedá premeniť na iný druh energie. Živé organizmy to na rozdiel od technických zariadení zásadne nedokážu. Stroj, ktorý využíva energiu chemických väzieb (napríklad spaľovací motor), ju najskôr premení na teplo a až potom na prácu: chemickú energiu paliva → teplý → práca (expanzia plynu vo valci a pohyb piestu). V živých organizmoch je možná iba táto schéma: chemická energia → Job.
Energia chemických väzieb v molekulách potravných látok je teda pre živočíšny organizmus prakticky jediným zdrojom energie a tepelnú energiu dokáže využiť len na udržanie telesnej teploty. Okrem toho teplo v dôsledku rýchleho rozptylu v prostredí nemôže byť v tele dlhodobo uchovávané. Ak sa v tele vyskytne nadmerné teplo, pre teplokrvné živočíchy sa to stáva vážnym problémom a niekedy dokonca ohrozuje ich život (pozri časť 11.3).
11.1. Zdroje energie a spôsoby jej premeny v organizme
Živý organizmus je otvorený energetický systém: energiu prijíma z okolia (takmer výlučne vo forme chemických väzieb), premieňa ju na teplo alebo prácu a v tejto forme ju vracia späť do okolia.
Zložky živín, ktoré vstupujú do krvi z gastrointestinálneho traktu (napríklad glukóza, mastné kyseliny alebo aminokyseliny), nie sú schopné priamo prenášať energiu svojich chemických väzieb na svojich konzumentov, napríklad draselno-sodnú pumpu alebo sval. aktín a myozín. Existuje univerzálny sprostredkovateľ medzi potravinovými „nosičmi energie“ a „spotrebiteľmi“ energie – adenozíntrifosfát (ATP). On je ten pravý priamy zdroj energie pre akékoľvek procesy v živých organizmoch
telo. Molekula ATP je kombináciou adenínu, ribózy a troch fosfátových skupín (obr. 11.1).
Väzby medzi zvyškami kyselín (fosfáty) obsahujú značné množstvo energie. Odštiepením koncového fosfátu pôsobením enzýmu ATPázy sa ATP premení na adenozíndifosfát (ADP). Tým sa uvoľní 7,3 kcal/mol energie. Energia chemických väzieb v molekulách potravy sa využíva na resyntézu ATP z ADP. Uvažujme tento proces s použitím glukózy ako príkladu (obr. 11.2).
Prvá fáza využitia glukózy je glykolýza. Počas tohto procesu sa najprv premení molekula glukózy na kyselina pyrohroznová (pyruvát), a zároveň poskytuje energiu na resyntézu ATP. Pyruvát sa potom prevedie na acetyl koenzým A - počiatočný produkt pre ďalšiu fázu recyklácie - Krebsov cyklus. Viacnásobné premeny látok, ktoré tvoria podstatu tohto cyklu, poskytujú dodatočnú energiu na resyntézu ATP a končia uvoľnením vodíkových iónov. Tretia etapa začína prenosom týchto iónov do dýchacieho reťazca - oxidačná fosforylácia, v dôsledku čoho vzniká aj ATP.
Celkovo všetky tri stupne recyklácie (glykolýza, Krebsov cyklus a oxidačná fosforylácia) tvoria proces tkanivové dýchanie. Zásadne dôležité je, aby prvý stupeň (glykolýza) prebiehal bez použitia kyslíka (anaeróbne dýchanie) a vedie k vytvoreniu iba dvoch molekúl ATP. Dve nasledujúce fázy (Krebsov cyklus a oxidačná fosforylácia) sa môžu vyskytnúť iba v kyslíkovom prostredí (aeróbne dýchanie).Úplné využitie jednej molekuly glukózy vedie k vzniku 38 molekúl ATP.
Existujú organizmy, ktoré nielenže nepotrebujú kyslík, ale v kyslíkovom (alebo vzdušnom) prostredí aj umierajú - povinných anaeróbov. Patria sem napríklad baktérie, ktoré spôsobujú plynovú gangrénu (Clostridium perfringes), tetanus (C. tetani), botulizmus (C. botulinum) atď.
U zvierat sú anaeróbne procesy pomocným typom dýchania. Napríklad pri intenzívnych a častých svalových kontrakciách (alebo pri statických kontrakciách) dodávka kyslíka krvou zaostáva za potrebami svalových buniek. V tomto čase dochádza k tvorbe ATP anaeróbne s akumuláciou pyruvátu, ktorý sa premieňa na kyselina mliečna (laktát). Rastúce kyslíkový dlh. Zastavenie alebo oslabenie svalovej práce odstraňuje rozpor medzi potrebou kyslíka tkaniva a možnosťami jeho dodávania sa laktát premieňa na pyruvát, ten sa buď cez štádium acetylkoenzýmu A oxiduje v Krebsovom cykle na oxid uhličitý, resp. glukoneogenézou sa mení na glukózu.
Podľa druhého termodynamického zákona dochádza k akejkoľvek transformácii energie z jedného typu na druhý s povinným vytvorením značného množstva tepla, ktoré sa potom rozptýli v okolitom priestore. Preto k syntéze ATP a prenosu energie z ATP k skutočným „spotrebiteľom energie“ dochádza so stratou približne polovice vo forme tepla. Zjednodušene môžeme tieto procesy znázorniť nasledovne (obr. 11.3).
Približne polovica chemickej energie obsiahnutej v potravinách sa okamžite premení na teplo a rozptýli sa v priestore, druhá polovica ide na tvorbu ATP. Pri následnom rozklade ATP sa polovica uvoľnenej energie opäť premení na teplo. Výsledkom je, že zviera a človek nemôžu minúť viac ako 1/4 všetkej energie spotrebovanej vo forme potravy na vykonávanie vonkajšej práce (napríklad beh alebo pohyb akýchkoľvek predmetov v priestore). Účinnosť vyšších zvierat a ľudí (asi 25 %) je teda niekoľkonásobne vyššia ako napríklad účinnosť parného stroja.
Všetka vnútorná práca (okrem procesov rastu a akumulácie tuku) sa rýchlo mení na teplo. Príklady: a) energia produkovaná srdcom sa premieňa na teplo v dôsledku odporu krvných ciev voči prietoku krvi; (b) žalúdok vykonáva sekréciu kyseliny chlorovodíkovej, pankreas vylučuje bikarbonátové ióny, v tenkom čreve tieto látky interagujú a v nich uložená energia sa mení na teplo.
Výsledky vonkajšej (užitočnej) práce vykonávanej zvieraťom alebo človekom sa tiež v konečnom dôsledku premenia na teplo: pohybom telies v priestore sa ohrieva vzduch, vzpriamené stavby sa zrútia, čím sa energia v nich uložená odovzdáva zemi a vzduchu vo forme tepla. Egyptské pyramídy sú vzácnym príkladom toho, ako energia svalovej kontrakcie, vynaložená pred takmer 5000 rokmi, stále čaká na nevyhnutnú premenu na teplo.
Rovnica energetickej bilancie:
E = A + H + S,
Kde E - celkové množstvo energie prijatej telom z potravy; A - externá (užitočná) práca; N - prenos tepla; S- uložená energia.
Straty energie močom, kožným mazom a inými sekrétmi sú extrémne malé a možno ich zanedbať.
Dýchací koeficient je 18,10:24,70 = 0,73.[...]
Dýchací koeficient nezostáva počas normálneho dozrievania ovocia konštantný. V premenopauzálnom štádiu je to približne 1 a dozrievaním dosahuje hodnoty 1,2... 1,5. Pri odchýlkach ±0,25 od jednej ešte nie sú v plodoch pozorované metabolické abnormality a len pri veľkých odchýlkach možno predpokladať fyziologické poruchy. Intenzita dýchania jednotlivých vrstiev tkaniva žiadneho plodu nie je rovnaká. V súlade s väčšou aktivitou enzýmov v koži je rýchlosť dýchania v nej mnohonásobne vyššia ako v parenchýmovom tkanive (Hulme a Rhodes, 1939). S poklesom obsahu kyslíka a zvýšením koncentrácie oxidu uhličitého v parenchýmových bunkách klesá intenzita dýchania so vzdialenosťou od šupky k jadru plodu [...].
Prístroj na stanovenie respiračného koeficientu, pinzeta, prúžky filtračného papiera, presýpacie hodiny na 2 minúty, sklenené poháre, pipety, sklenené tyčinky, 250 ml kužeľové banky.[...]
Prístroj na stanovenie respiračného koeficientu pozostáva z veľkej skúmavky s tesne priliehajúcou gumenou zátkou, do ktorej sa vkladá do pravého uhla zahnutá meracia trubica so stupnicou milimetrového papiera.[...]
Spotreba kyslíka a koeficient jeho využitia boli konštantné, keď sa p02 znížilo na 60 a 20 % pôvodného (v závislosti od prietoku). Pri koncentráciách kyslíka mierne nad kritickou úrovňou sa maximálny objem ventilácie udržiaval dlhodobo (niekoľko hodín). Objem ventilácie sa zvýšil 5,5-krát, ale na rozdiel od kaprov sa znížil od 22% úrovne nasýtenia vody kyslíkom. Autori sa domnievajú, že zníženie objemu ventilácie u rýb pri extrémnej hypoxii je dôsledkom nedostatku kyslíka v dýchacích svaloch. Pomer frekvencie dýchania a srdcovej frekvencie bol 1,4 normálne a 4,2 s nedostatkom kyslíka.
Úvodné vysvetlivky. Výhody metódy: vysoká citlivosť, ktorá umožňuje pracovať s malými vzorkami experimentálneho materiálu; schopnosť sledovať dynamiku výmeny plynov a súčasne zohľadňovať výmenu plynov 02 a C02, čo umožňuje nastaviť koeficient dýchania.[...]
Preto hodnota pH v oxyteiku klesá na takmer 6,0, kým v prevzdušňovacej nádrži pH>7D Pri maximálnom zaťažení je príkon pre oxytank vrátane výkonu zariadenia na výrobu kyslíka 1,3 m3/ (hp-h). a výkonového prevzdušňovača (obr. 26.9), by mal byť menší ako výkon prevzdušňovača pre prevzdušňovaciu nádrž. Vysvetľuje to vysoká koncentrácia kyslíka (nad 60 %) vo všetkých stupňoch kyslíkovej nádrže.[...]
Dynamika uvoľňovania oxidu uhličitého (С?СО2), absorpcia kyslíka ([...]
Morské a sladkovodné ryby mali za týchto experimentálnych podmienok približne rovnaký respiračný koeficient (RQ). Nevýhodou týchto údajov je, že autor zobral na porovnanie zlatú rybku, ktorá vo všeobecnosti spotrebováva málo kyslíka a len ťažko môže slúžiť ako etalón porovnania.[...]
V súvislosti s výmenou plynov zimujúceho hmyzu treba povedať, že klesá aj respiračný koeficient1. Napríklad Dreyer (1932) zistil, že v aktívnom stave mravca Formica ulkei Emery bol koeficient dýchania 0,874; keď sa mravce stali neaktívnymi pred hibernáciou, respiračný koeficient sa znížil na 0,782 a počas obdobia hibernácie pokles dosiahol 0,509-0,504. Zemiakový chrobák z Colorada Leptinotarsa decemlineata Say. v období zimovania respiračný koeficient klesá na 0,492-0,596, zatiaľ čo v lete je to 0,819-0,822 (Ushatinskaya, 1957). Vysvetľuje to skutočnosť, že v aktívnom stave sa hmyz živí najmä bielkovinovými a uhľohydrátovými potravinami, zatiaľ čo v hibernácii sa spotrebuje hlavne tuk, ktorý vyžaduje menej kyslíka na oxidáciu.
V uzavretých nádobách určených pre tlak v GP RK. d = 1962 Pa (200 mm vodného stĺpca), pri vysokých rýchlostiach obrátky môže byť doba nečinnosti nádrže s „mŕtvym“ zvyškom pred začatím plnenia taká krátka, že sa dýchací ventil nestihne otvoriť na „výdych“ “. Potom nedochádza k stratám zo „spätného výdychu“.[...]
Pre pochopenie biochemických procesov prebiehajúcich v tele má veľký význam hodnota respiračného koeficientu. Respiračný koeficient (RC) je pomer vydychovanej kyseliny uhličitej a spotrebovaného kyslíka.[...]
Na posúdenie vplyvu teploty na akýkoľvek proces zvyčajne pracujú na hodnote teplotného koeficientu. Teplotný koeficient (t>ω) dýchacieho procesu závisí od druhu rastliny a teplotných gradácií. So zvýšením teploty z 5 na 15 °C sa teda 0 ω môže zvýšiť na 3, zatiaľ čo zvýšenie teploty z 30 na 40 °C zvyšuje intenzitu dýchania menej výrazne (ω asi 1,5). Fáza vývoja rastlín má veľký význam. Podľa B., A. Rubina sú v každej fáze vývoja rastliny pre proces dýchania najpriaznivejšie teploty, na pozadí ktorých táto fáza zvyčajne prebieha Zmena optimálnych teplôt pri dýchaní rastlín v závislosti od fázy o ich vývoj je spôsobený tým, že v procese ontogenézy menia dýchacie výmenné cesty. Medzitým sú rôzne teploty najpriaznivejšie pre rôzne enzýmové systémy. V tomto smere je zaujímavé, že v neskorších fázach vývoja rastlín sa pozorujú prípady, kedy flavíndehydrogenázy pôsobia ako finálne oxidázy, odovzdávajúce vodík priamo vzdušnému kyslíku.[...]
Všetky skúmané ryby v zajatí spotrebujú menej kyslíka ako v prirodzených podmienkach. Mierne zvýšenie respiračného koeficientu u rýb chovaných v akváriách naznačuje zmenu kvalitatívnej stránky metabolizmu smerom k väčšej účasti sacharidov a bielkovín v ňom. Autor to vysvetľuje horším kyslíkovým režimom akvária v porovnaní s prírodnými podmienkami; Okrem toho sú ryby v akváriu neaktívne.[...]
Na zníženie emisií výparov škodlivých látok sa používajú aj reflektorové kotúče, inštalované pod montážnou rúrkou dýchacieho ventilu. Pri vysokej rýchlosti obrátky atmosférických nádrží môže účinnosť reflektorových kotúčov dosiahnuť 20-30%.[...]
K opätovnému nasýteniu plynovej komory môže dôjsť po naplnení, ak plynový priestor nebol úplne nasýtený parami. V tomto prípade sa dýchací ventil po naplnení nádoby neuzavrie a okamžite začne ďalší výdych. Tento jav sa vyskytuje v nádržiach, ktoré majú vysoký obratový pomer alebo sú čiastočne naplnené, nie do maximálnej výšky plnenia, ako aj v nádržiach s pomalými procesmi saturácie hydraulickej kvapaliny (nádrže s pontónmi a zapustené). Sýtosť GP je typická najmä pre nádrže, ktoré sa plnia prvýkrát po vyčistení a odvetraní. Tento typ straty sa niekedy nazýva straty z nasýtenia alebo presýtenia GP.[...]
Pre známe u0 Acjcs možno tiež určiť z grafov podobných tým, ktoré sú znázornené na obr. 14. Metódy na výpočet strát poskytujú podobné grafy pre typické nádrže RVS, rôzne typy dýchacích ventilov a ich množstvá. Hodnota Ac/cs znamená nárast koncentrácie v čerpacej stanici počas celkovej doby odstávky (tp) a plnenia zásobníka (te), t.j. t = t„ + t3; určí sa približne z grafov (pozri obr. 3). Pri použití vzorca (!9) je potrebné mať na pamäti, že pri úplnom nasýtení GP ccp/cs = 1 a že čas na úplné nasýtenie GP pozemných nádrží je obmedzený na 2-4 dni ( v závislosti od poveternostných podmienok a iných podmienok) a graf na " Obr. 3 približný. Preto po získaní hodnôt ccp/cs>l zo vzorca (19), čo znamená začiatok úplného nasýtenia dodávky plynu pred koniec odstávky alebo koniec plnenia nádrže, je potrebné nahradiť ccp/cs = 1.[ ..]
Vyhodnoťme kvantitatívne vzťahy medzi týmito dvoma prúdmi plynu. Po prvé, pomer objemu uvoľneného oxidu uhličitého k objemu spotrebovaného kyslíka (respiračný koeficient) pre väčšinu odpadových vôd a aktivovaného kalu je menší ako jedna. Po druhé, objemové koeficienty prenosu hmoty pre kyslík a oxid uhličitý sú blízko seba. Po tretie, fázová rovnovážna konštanta oxidu uhličitého je takmer 30-krát menšia ako konštanta kyslíka. Po štvrté, oxid uhličitý nie je prítomný len v kalovej zmesi v rozpustenom stave, ale vstupuje aj do chemickej interakcie s vodou.[...]
Pri porovnaní oboch typov dýchania je markantný nerovnaký pomer absorpcie kyslíka k uvoľňovaniu oxidu uhličitého. Pomer CO2/O2 sa označuje ako respiračný koeficient KO.[...]
Ak sa pri dýchaní oxidujú organické látky s relatívne vyšším obsahom kyslíka ako v sacharidoch, napríklad organické kyseliny - šťaveľová, vínna a ich soli, potom bude respiračný koeficient výrazne väčší ako 1. Bude tiež väčší ako 1 v prípade keď sa časť kyslíka, používaného na mikrobiálne dýchanie, odoberá zo sacharidov; alebo pri dýchaní tých kvasiniek, v ktorých alkoholová fermentácia prebieha súčasne s aeróbnym dýchaním. Ak sa popri aeróbnom dýchaní vyskytnú aj iné procesy, pri ktorých sa používa dodatočný kyslík, potom bude koeficient dýchania menší ako 1. Bude tiež menší ako 1, keď látky s relatívne nízkym obsahom kyslíka, ako sú bielkoviny, uhľovodíky atď. ., sú oxidované počas dýchacieho procesu Následne, ak poznáte hodnotu respiračného koeficientu, môžete určiť, ktoré látky sú oxidované počas dýchania.
Najvšeobecnejším ukazovateľom rýchlosti oxidácie je rýchlosť dýchania, ktorú možno posúdiť podľa absorpcie kyslíka, uvoľňovania oxidu uhličitého a oxidácie organických látok. Ďalšie ukazovatele respiračného metabolizmu: hodnota respiračného koeficientu, pomer glykolytických a pentózofosfátových ciest štiepenia cukrov, aktivita redoxných enzýmov. Energetickú účinnosť dýchania možno posudzovať podľa intenzity oxidačnej fosforylácie mitochondrií.[...]
Trendy uvedené pre jablká Cox Orange týkajúce sa vplyvu koncentrácií kyslíka a oxidu uhličitého vo vzduchu v komore platia pre všetky ostatné odrody jabĺk, s výnimkou prípadov, keď koeficient dýchania rastie s klesajúcou teplotou výraznejšie.
Hodnota DC závisí od iných dôvodov. V niektorých tkanivách v dôsledku ťažkého prístupu kyslíka spolu s aeróbnym dýchaním dochádza k anaeróbnemu dýchaniu, ktoré nie je sprevádzané absorpciou kyslíka, čo vedie k zvýšeniu hodnoty DC. Hodnotu koeficientu určuje aj úplnosť oxidácie dýchacieho substrátu. Ak sa v tkanivách okrem finálnych produktov hromadia aj menej oxidované zlúčeniny (organické kyseliny), potom DC[...]
Kvantitatívne stanovenia závislosti výmeny plynov v rybách na teplote boli vykonané mnohými výskumníkmi. Vo väčšine prípadov sa štúdium tejto problematiky obmedzilo predovšetkým na kvantitatívnu stránku dýchania – veľkosť dýchacieho rytmu, množstvo spotreby kyslíka a následne výpočet teplotných koeficientov pri rôznych teplotách.[...]
Na zníženie strát odparovaním a znečistením ovzdušia sú benzínové nádrže vybavené plynovým potrubím spájajúcim vzduchové priestory nádrží, v ktorých sú skladované produkty rovnakej značky, a je inštalovaný spoločný dýchací ventil. Vyššie popísané „veľké a malé dýchanie“, vetranie plynového priestoru, tiež spôsobuje znečistenie ovzdušia pri skladovaní ropných produktov v poľnohospodárskych zariadeniach, keďže pri obratovom pomere tankovej farmy 4-6 je obrat zásob paliva 10- 20, čo znamená pokles pomerového využitia nádrží 0,4-0,6. Aby sa zabránilo znečisteniu ovzdušia, sú sklady ropy vybavené čistiacimi zariadeniami a lapačmi benzínu a oleja.[...]
Doteraz získané údaje ukazujú, že extrémne teploty spôsobujú inhibíciu fyziologického systému, najmä transportu plynov v rybách. Súčasne sa rozvíja bradykardia, zvyšuje sa arytmia, klesá spotreba kyslíka a miera jeho využitia. Po týchto zmenách vo fungovaní kardiorespiračného aparátu postupne zaniká ventilácia žiabier a v poslednom rade prestáva fungovať myokard. Jedným z dôvodov úhynu rýb v dôsledku prehriatia je zrejme anoxia dýchacích svalov a celkový nedostatok kyslíka. Zvýšenie teploty vedie k zrýchleniu využitia kyslíka a v dôsledku toho k poklesu jeho napätia v dorzálnej aorte, čo zase slúži ako signál pre zvýšenú ventiláciu žiabier.[...]
Pred použitím modelu je potrebné skontrolovať jeho kinetické parametre. Validáciu modelu čistého kyslíkového systému na čistenie domových a priemyselných odpadových vôd vykonali Muller et al.(1) Validácia modelu na čistenie domových odpadových vôd používala respiračný koeficient R.C 1,0, kým pre priemyselné odpadové vody je to 0,85 resp. dokonca 0,60 bolo urobené celkom nedávno pri štúdiu odpadových vôd z celulózky a papierne (obr. 26.6). a bola zaznamenaná nižšia požiadavka na rast mikroorganizmov, ako sa tradične pozoruje v biologických systémoch [...].
Na vyriešenie otázky podstaty vplyvu teploty na metabolizmus rýb je potrebné poznať nielen mieru zvýšenia alebo zníženia metabolizmu so zmenou teploty, ale aj kvalitatívne zmeny v jednotlivých väzbách, ktoré tvoria metabolizmus. Kvalitatívnu stránku metabolizmu možno do určitej miery charakterizovať takými koeficientmi ako respiračný a amoniakálny (pomer uvoľneného amoniaku ako konečného produktu metabolizmu dusíka k spotrebovanému kyslíku) (obr. 89).[...]
Z vyššie uvedenej rovnice (4) vyplýva, že pomer konštánt pre 02 a CO2 sa rovná 1,15, t. j. použitie techniky merania bilancie CO2 by zrejme umožnilo vykonávať pozorovania pri mierne vyšších hodnotách 2. a zodpovedajúco vyššie rýchlosti prúdenia. Táto zjavná výhoda však zmizne, ak predpokladáme, že koeficient dýchania je menší ako 1. Okrem toho, ako ukázal Talling 32], presnosť stanovenia CO2 v prírodných vodách nemôže byť lepšia ako ± 1 µmol/l (0,044 mg/l), a kyslík - ±0,3 umol/l (0,01 mg/l). V dôsledku toho, aj keď vezmeme respiračný koeficient rovný 1, presnosť bilančnej metódy, založenej na zohľadnení bilancie kyslíka, sa ukáže byť najmenej trikrát vyššia ako pri stanovení oxidu uhličitého.[...]
V našich štúdiách bola použitá morfofyziologická metóda s niektorými doplnkami. To umožnilo s dostatočnou presnosťou (±3,5 %) určiť množstvo absorbovaného kyslíka, uvoľneného oxidu uhličitého a koeficient dýchania (RQ) na celých sadeniciach starých 10-12 dní a listoch rastlín z poľných pokusov. Princíp tejto techniky spočíva v tom, že rastliny umiestnené v uzavretej nádobe (špeciálne navrhnutá plynová pipeta) s atmosférickým vzduchom menia zloženie vzduchu v dôsledku dýchania. Keď teda poznáme objem nádoby a určíme percentuálne zloženie vzduchu na začiatku a na konci experimentu, nie je ťažké vypočítať množstvo absorbovaného a uvoľneného CO2 rastlinami [...].
Rôzne rastlinné orgány a tkanivá sa značne líšia v podmienkach zásobovania kyslíkom. V liste kyslík voľne prúdi takmer do každej bunky. Šťavnaté ovocie, korene, hľuzy sú veľmi zle vetrané; sú slabo priepustné pre plyny, nielen pre kyslík, ale aj pre oxid uhličitý. Prirodzene, v týchto orgánoch sa dýchací proces posúva na anaeróbnu stranu a dýchací koeficient sa zvyšuje. V meristematických tkanivách sa pozoruje zvýšenie respiračného koeficientu a posun procesu dýchania na anaeróbnu stranu. Rôzne orgány sa teda vyznačujú nielen rôznou intenzitou, ale aj nerovnakou kvalitou dýchacieho procesu.[...]
Otázka látok používaných v procese dýchania je už dlho témou pre fyziológov. Aj v prácach I.P. Borodina sa ukázalo, že intenzita dýchacieho procesu je priamo úmerná obsahu sacharidov v rastlinných tkanivách. To dalo dôvod predpokladať, že sacharidy sú hlavnou látkou spotrebovanou počas dýchania. Pri objasňovaní tejto problematiky má veľký význam určenie respiračného koeficientu. Respiračný koeficient je objemový alebo molárny pomer CO2 uvoľneného počas dýchania k CO2 absorbovanému počas rovnakého časového obdobia Pri normálnom prístupe kyslíka závisí hodnota respiračného koeficientu od substrátu dýchania. Ak sa v procese dýchania používajú sacharidy, potom proces prebieha podľa rovnice CeH) 2O5 + 6O2 = 6CO2 + 6H2O, v tomto prípade je koeficient dýchania rovný jednej = 1. Ak sa však pri dýchaní rozkladá viac oxidovaných zlúčenín, ako sú organické kyseliny, absorpcia kyslíka sa znižuje a koeficient dýchania je väčší ako jeden. Keď sa počas dýchania oxiduje viac redukovaných zlúčenín, ako sú tuky alebo bielkoviny, vyžaduje sa viac kyslíka a koeficient dýchania sa stáva menším ako jedna.[...]
Takže najjednoduchší proces aeróbneho dýchania je znázornený v nasledujúcej forme. Molekulárny kyslík spotrebovaný pri dýchaní sa využíva najmä na viazanie vodíka vznikajúceho pri oxidácii substrátu. Vodík zo substrátu sa prenáša na kyslík prostredníctvom série medzireakcií, ktoré prebiehajú postupne za účasti enzýmov a nosičov. Takzvaný respiračný koeficient poskytuje určitú predstavu o povahe dýchacieho procesu. Rozumie sa to ako pomer objemu uvoľneného oxidu uhličitého k objemu kyslíka absorbovaného počas dýchania (C02:02).[...]
Výkonnosť kardiorespiračného aparátu rýb, jeho rezervné schopnosti a labilita frekvenčných a amplitúdových parametrov závisia od druhu a ekologických vlastností rýb. Pri zvýšení teploty o rovnakú hodnotu (z 5 na 20°C) sa dychová frekvencia zubáča zvýšila z 25 na 50 za minútu, u šťuky zo 46 na 75 a u ide zo 63 na 112 za minútu. Spotreba kyslíka sa zvyšuje súbežne so zvyšujúcou sa frekvenciou, ale nie hĺbkou dýchania. Najväčší počet dýchacích pohybov na prečerpanie jednotkového objemu vody vyprodukuje ide o mobilný ide a najmenej menej aktívny oxyfilný zubáč, čo pozitívne koreluje s intenzitou výmeny plynov u skúmaného druhu. Pomer maximálneho objemu ventilácie a zodpovedajúceho koeficientu využitia kyslíka podľa autorov určuje maximálne energetické možnosti organizmu. V pokoji bola najväčšia intenzita výmeny plynov a objem ventilácie u oxyfilných zubáčov a pri funkčnej záťaži (motorická aktivita, hypoxia) - v ide. Pri nízkych teplotách bolo zvýšenie objemu ventilácie v ide v reakcii na hypoxiu väčšie ako pri vysokých teplotách, konkrétne: 20-násobné pri 5 °C a 8-násobné pri 20 °C. V Orthologus thioglossy sa pri hypoxii (40% saturácia) objem vody čerpanej cez žiabre mení v menšej miere: pri 12°C sa zväčší 5-krát a pri 28°C - 4,3-krát.[...]
Ukazovatele metabolizmu uhľohydrátov počas adaptívnej exogénnej hypoxie, t.j. počas mierneho až stredného nedostatku kyslíka v prostredí, boli oveľa menej úplne študované. Obmedzené dostupné experimentálne údaje však ukazujú, že v tomto prípade dochádza k zvýšenému využitiu glykogénu vo svaloch, zvýšeniu kyseliny mliečnej a cukru v krvi. Ako by sa dalo očakávať, úroveň nasýtenia vody kyslíkom, pri ktorej dochádza k týmto posunom, sa u jednotlivých druhov líši. Napríklad u lamprey bola zaznamenaná hyperglykémia, keď obsah kyslíka klesol len o 20 % pôvodnej úrovne a u 1 abeo karepvka zostala koncentrácia cukru v krvi konštantne nízka aj pri 40 % nasýtení vody kyslíkom a len ďalší zníženie saturácie viedlo k rýchlemu zvýšeniu hladiny cukru v krvi. Pri hypoxii u lieňov bolo zaznamenané zvýšenie hladiny cukru v krvi a kyseliny mliečnej. Podobná reakcia na hypoxiu bola zaznamenaná pri kanálových sumcoch. V prvej z týchto štúdií sa pri 50% nasýtení vody kyslíkom u rýb zistilo zvýšenie obsahu kyseliny mliečnej, ktoré pokračovalo v prvej hodine normoxie, teda po návrate rýb do normálnych kyslíkových podmienok. Obnova biochemických parametrov do normálu nastala v priebehu 2-6 hodín a zvýšenie obsahu laktátu a respiračného koeficientu z 0,8 na 2,0 indikovalo zvýšenie anaeróbnej glykolýzy.
Respiračný kvocient (RC) je pomer objemu uvoľneného oxidu uhličitého k objemu kyslíka absorbovaného za určitý čas. Ak sa počas metabolického procesu v tele oxidujú iba uhľohydráty, potom sa respiračný koeficient bude rovnať 1. To možno vidieť z nasledujúceho vzorca:
V dôsledku toho na vytvorenie jednej molekuly C02 počas metabolizmu sacharidov je potrebná jedna molekula O2. Pretože podľa Avogadro-Gerardovho zákona rovnaký počet molekúl pri rovnakej teplote a tlaku zaberá rovnaký objem. Preto sa respiračný koeficient pre oxidáciu uhľohydrátov bude rovnať 1:
![]()
Pre tuky to bude:
![]()
Na oxidáciu jednej molekuly tuku je potrebných 81,5 molekúl kyslíka a na oxidáciu 1 gramu molekuly tuku je potrebných 81,5 x 22,4 litra kyslíka, teda 1825,6 litra O2, kde 22,4 je objem jedného gramu molekuly v litroch. . Gram molekuly tuku sa rovná 890 g, potom oxiduje 1 liter kyslíka ![]() 487 g tuku. 1 g tuku po úplnej oxidácii uvoľní 38,945 kJ (9,3 kcal)* a 0,487 dáva 18,551 kJ. Preto sa kalorický ekvivalent 1 litra kyslíka s respiračným koeficientom 0,7 bude rovnať 18,551 kJ. Za normálnych podmienok je respiračný koeficient medzi 1 a 0,7. Pri DC 0,7 sa v tele oxidujú tuky a kalorický ekvivalent, čiže kalorická hodnota 1 litra kyslíka je 18,551 kJ a pri DC 1 je to 21,135.
487 g tuku. 1 g tuku po úplnej oxidácii uvoľní 38,945 kJ (9,3 kcal)* a 0,487 dáva 18,551 kJ. Preto sa kalorický ekvivalent 1 litra kyslíka s respiračným koeficientom 0,7 bude rovnať 18,551 kJ. Za normálnych podmienok je respiračný koeficient medzi 1 a 0,7. Pri DC 0,7 sa v tele oxidujú tuky a kalorický ekvivalent, čiže kalorická hodnota 1 litra kyslíka je 18,551 kJ a pri DC 1 je to 21,135.