Определение дыхательного коэффициента (ДК) растений. Определение дыхательного коэффициента Дыхательный коэффициент его значение величина
дыхательный коэффициент (ДК)
отношение объема выделенного через легкие углекислого газа к объему поглощенного за это же время кислорода; величина Д. к. при пребывании исследуемого в покое зависит от вида окисленных в организме пищевых веществ.
Энциклопедический словарь, 1998 г.
дыхательный коэффициент
отношение объема углекислого газа, выделенного за определенное время при дыхании, к объему поглощенного за то же время кислорода. Характеризует особенности газообмена и обмена веществ у животных и растений. У здорового человека равен примерно 0,85.
Дыхательный коэффициент
отношение объёма выделяемого из организма углекислого газа к объёму поглощаемого за то же время кислорода. Обозначается:
Определение ДК важно для исследования особенностей газообмена и обмена веществ у животных и растительных организмов. При окислении в организме углеводов и полном доступе кислорода ДК равен 1, жиров ≈ 0,7, белков ≈ 0,8. У здорового человека в покое ДК равен 0,85 ╠ 0,1; при умеренной работе, а также у животных, питающихся преимущественно растительной пищей, приближается к 1. У человека при очень длительной работе, голодании, у плотоядных животных (хищников), а также при спячке, когда из-за ограниченности запасов углеводов в организме усиливается диссимиляция жиров, ДК составляет около 0,7. ДК превышает 1 при интенсивном отложении в организме жиров, образующихся из поступающих с пищей углеводов (например, у человека при восстановлении нормального веса после голодания, после длительных заболеваний, а также у животных при откорме). До 2 ДК возрастает при усиленной работе и гипервентиляции лёгких, когда из организма выделяется дополнительно СО2, находившийся в связанном состоянии. Ещё больших величин ДК достигает у анаэробов, у которых большая часть выделяемого CO2 образуется путём бескислородного окисления (брожения). ДК ниже 0,7 бывает при заболеваниях, связанных с нарушениями обмена веществ, после тяжёлой физической работы.
У растений ДК зависит от химической природы дыхательного субстрата, содержания CO2 и O2 в атмосфере и др. факторов, характеризуя, т. о., специфику и условия дыхания. При использовании клеткой для дыхания углеводов (проростки злаков) ДК равен примерно 1, жиров и белков (прорастающие семена масличных и бобовых) ≈ 0,4≈0,7. При недостатке О2 и затруднённом его доступе (семена с твёрдой оболочкой) ДК равен 2≈3 и более; высокий ДК характерен также для клеток точек роста.
(ДК) это отношение объема выделенного в процессе дыхания углекислого газа к объему поглощенного кислорода.
Величина дыхательного коэффициента растений
Величина ДК указывает как на характер окисляемого в процессе дыхания материала, так и на тип дыхания; она может быть равна единице, больше или меньше ее. При окислении углеводов объемы обмениваемых газов углекислоты и кислорода равны и отношение С0 2: 0 2 равно единице. В данном случае потребляемый при дыхании кислород идет только на окисление углерода до углекислоты, потому что соотношение водорода и кислорода в молекуле глюкозы таково, что для окисления водорода до воды кислорода достаточно в самой молекуле сахара. При окислении ряда органических кислот дыхательный коэффициент растений больше единицы. Так, щавелевая кислота - соединение, более богатое кислородом, чем углеводы. Кислорода, имеющегося в молекуле, не только достаточно для окисления водорода до воды, но часть его остается и для окисления углерода; поэтому для полного окисления двух молекул щавелевой кислоты достаточно одной молекулы кислорода: 2С 2 Н 2 О 4 + О 2 → 4СО 2 + 2Н 2 О, ДК (4СО 2: О 2) в этом случае равен 4. В тех случаях, когда растение дышит за счет белков или жиров, в молекуле которых много водорода и углерода и мало кислорода, ДК меньше единицы, так как для окисления всего углерода и водорода, находящегося в этих соединениях, необходимо поглотить большое количество кислорода. При окислении стеариновой кислоты реакция окисления пойдет следующим образом: С 18 Н 26 О 2 + 26О 2 → 18СО 2 + 18Н 2 О. ДК (18СО 2: 26О 2) равен 0,69. Таким образом, в случае окисления углеводов ДК равен единице, органических кислот - больше единицы, белков и жиров - меньше единицы.
Тепловой эффект при дыхании растений
Тепловой эффект будет иметь значение, обратное величине ДК: максимальный тепловой эффект будет при окислении жиров, потому что они наиболее восстановленные соединения. Зависимость величины ДК от характера дыхательного материала наблюдается только в том случае, когда в окружающей среде и тканях растения достаточно кислорода. Однако при окислении одного и того же дыхательного материала, но при недостатке кислорода в окружающей среде и тканях растений величины ДК также могут изменяться. Если кислорода мало, то при окисление идет не до конца и кроме углекислого газа и воды образуются органические кислоты, которые более окислены, чем углеводы. В этом случае ДК будет меньше единицы, так как часть поглощенного кислорода останется в молекулах образованных органических кислот, углекислоты же выделится меньше. Меньше выделится и энергии, так как часть ее сохранится в органических кислотах.1. Какой процесс обеспечивает освобождение энергии в организме? В чем его сущность?
Диссимиляция (катаболизм), т. е. распад клеточных структур и соединений организма с выделением энергии и продуктов распада.
2. Какие питательные вещества служат источником энергии в организме?
Углеводы, жиры и белки.
3. Назовите основные методы определения количества энергии в навеске продукта.
Физическая калориметрия; физико-химические методики определения количества питательных веществ в навеске с последующим расчетом содержащейся в ней энергии; по таблицам.
4. Опишите сущность способа физической калориметрии.
В калориметре сжигают навеску продукта, а затем по степени нагревания воды и материала калориметра рассчитывают выделившуюся энергию.
5. Напишите формулу расчета количества тепла, выделившегося при сгорании продукта в калориметре. Расшифруйте ее обозначения.
Q = MвСв (t 2 - t 1) + MкСк (t 2 - t 1) - Qо,
где Q - количество тепла, М – масса (в – воды, к – калориметра), (t 2 - t 1) – разность температур воды и калориметра после и до сжигания навески, С – удельная теплоемкость, Qо -количество тепла, образуемое окислителем.
6. Что называют физическим и физиологическим калорическими коэффициентами питательного вещества?
Количество тепла, освобождаемое при сгорании 1 г вещества в калориметре и в организме соответственно.
7. Сколько тепла освобождается при сгорании в калориметре 1 г белков, жиров и углеводов?
1г белков – 5, 85 ккал (24, 6 кДж), 1г жиров – 9, 3 ккал (38, 9 кДж), 1г углеводов – 4, 1 ккал (17, 2 кДж).
8. Сформулируйте закон термодинамики Гесса, на основании которого рассчитывают приход энергии в организм по количеству усвоенных белков, жиров и углеводов.
Термодинамический эффект зависит только от теплосодержания начальных и конечных продуктов реакции и не зависит от промежуточных превращений этих веществ.
9. Сколько тепла освобождается при окислении в организме 1 г белков, 1 г жиров и 1 г углеводов?
1 г белков – 4, 1 ккал (17, 2 кДж), 1 г жиров – 9, 3 ккал (38, 9 кДж), 1 г углеводов – 4, 1 ккал (17, 2 кДж).
10. Объясните причину различия физического и физиологического калорических коэффициентов для белков. В каком случае он больше?
В калориметре (физический коэффициент) белок распадается до конечных продуктов – СО 2 , Н 2 О и NН 3 c освобождением всей заключенной в них энергии. В организме (физиологический коэффициент) белки распадаются до СО 2 , Н 2 О, мочевины и др. веществ белкового обмена, которые содержат энергию и выводятся с мочой.
Определяют содержание белков, жиров и углеводов в продуктах питания, умножают их количество на соответствующие физиологические калорические коэффициенты, суммируют и из суммы вычитают 10%, что не усваивается в пищеварительном тракте (потери с калом).
12. Рассчитайте (в ккал и кДж) приход энергии при поступлении в организм с пищей по 10 г белков, жиров и углеводов.
Q = 4,110 + 9,310 + 4,110 = 175 ккал. (175 ккал – 17,5 ккал) х 4,2 кДж, где 17,5 ккал – энергия неусвоенных питательных веществ (потери с калом – около 10%). Итого: 157,5 ккал (661,5 кДж).
Калориметрия: прямая (метод Этуотера – Бенедикта); непрямая, или косвенная (методы Крога, Шатерникова, Дугласа – Холдена).
14. На чем основан принцип прямой калориметрии?
На непосредственном измерении количества тепла, выделенного организмом.
15. Опишите кратко устройство и принцип работы камеры Этуотера – Бенедикта.
Камера, в которую помещают испытуемого, термически изолирована от окружающей среды, ее стенки не поглощают тепло, внутри них находятся радиаторы, через которые течет вода. По степени нагрева определенной массы воды рассчитывают количество тепла, израсходованного организмом.
16. На чем основан принцип непрямой (косвенной) калориметрии?
На расчете количества выделившейся энергии по данным газообмена (поглощенный О 2 и выделившийся СО 2 за сутки).
17. Почему количество выделяемой организмом энергии можно рассчитать по показателям газообмена?
Потому, что количество потребленного организмом О 2 и выделенного СО 2 точно соответствует количеству окисленных белков, жиров и углеводов, а значит, и израсходованной организмом энергии.
18. Какие коэффициенты используются для расчета расхода энергии методом непрямой калориметрии?
Дыхательный коэффициент и калорический эквивалент кислорода.
19. Что называют дыхательным коэффициентом?
Отношение объема выделенного организмом углекислого газа к объему потребленного за это же время кислорода.
20. Рассчитайте дыхательный коэффициент (ДК), если известно, что во вдыхаемом воздухе содержится 17% кислорода и 4% углекислого газа.
Так как в атмосферном воздухе содержится 21% О 2 , процент поглощенного кислорода составляет 21% – 17%, т. е. 4 %. СО 2 в выдыхаемом воздухе также составляет 4%. Отсюда
21. От чего зависит величина дыхательного коэффициента?
22. Чему равен дыхательный коэффициент при окислении в организме до конечных продуктов белков, жиров и углеводов?
При окислении белков – 0,8, жиров – 0,7, углеводов – 1, 0.
23. Почему дыхательный коэффициент для жиров и белков ниже, чем для углеводов?
На окисление белков и жиров расходуется больше О 2 , так как они содержат меньше внутримолекулярного кислорода, чем углеводы.
24. К какой величине приближается дыхательный коэффициент у человека в начале интенсивной физической работы? Почему?
К единице, потому что источником энергии в этом случае являются преимущественно углеводы.
25. Почему в первые минуты после интенсивной и длительной физической работы дыхательный коэффициент у человека больше единицы?
Потому что СО 2 выделяется больше, чем потребляется О 2 , так как молочная кислота, накопившаяся в мышцах, поступает в кровь и вытесняет СО 2 из бикарбонатов.
26. Что называют калорическим эквивалентом кислорода?
Количество тепла, освобождаемое организмом при потреблении 1л О 2 .
27. От чего зависит величина калорического эквивалента кислорода?
От соотношения белков, жиров и углеводов, окисляющихся в организме.
28. Чему равен калорический эквивалент кислорода при окислении в организме (в процессе диссимиляции) белков, жиров и углеводов?
Для белков – 4, 48 ккал (18, 8 кДж), для жиров – 4, 69 ккал (19, 6 кДж), для углеводов – 5, 05 ккал (21, 1 кДж).
29. Опишите кратко ход определения расхода энергии по способу Дугласа – Холдена (полный газовый анализ).
В течение нескольких минут испытуемый вдыхает атмосферный воздух, а выдыхаемый воздух собирают в специальный мешок, измеряют его количество и проводят анализ газов с целью определения объема потребленного кислорода и выделившегося СО 2 . Рассчитывают дыхательный коэффициент, с помощью которого по таблице находят соответствующий калорический эквивалент О 2 , который затем умножают на объем О 2 , потребленного за данный промежуток времени.
30. Опишите кратко метод М. Н. Шатерникова для определения расхода энергии у животных в эксперименте.
Животное помещают в камеру, в которую поступает кислород по мере его расходования. Выделяющийся при дыхании СО 2 поглощается щелочью. Расчет выделенной энергии осуществляется по количеству потребленного О 2 и усредненному калорическому эквиваленту О 2: 4,9 ккал (20,6 кДж).
31. Рассчитайте расход энергии за 1 минуту, если известно, что испытуемый потребил 300 мл О 2 . Дыхательный коэффициент равен 1,0.
ДК=1,0, ему соответствует калорический эквивалент кислорода, равный 5,05 ккал (21,12 кДж). Следовательно, расход энергии за минуту = 5,05 ккал х 0,3 = 1,5 ккал (6,3 кДж).
32. Опишите кратко ход определения расхода энергии по способу Крога у человека (неполный газовый анализ).
Испытуемый вдыхает кислород из мешка метаболиметра, выдыхаемый воздух возвращается в тот же мешок, предварительно пройдя через поглотитель СО 2 . По показаниям метаболиметра определяют расход О 2 и умножают на калорический эквивалент кислорода 4,86 ккал (20,36 кДж).
33. Назовите основные различия в расчете расхода энергии по способам Дугласа – Холдена и Крога.
Метод Дугласа – Холдена предполагает расчет расхода энергии по данным полного газового анализа; метод Крога – только по объему потребленного кислорода с использованием калорического эквивалента кислорода, характерного для условий основного обмена.
34. Что называют основным обменом?
Минимальный расход энергии, обеспечивающий гомеостазис в стандартных условиях: при бодрствовании, максимальном мышечном и эмоциональном покое, натощак (12 – 16 часов без еды), при температуре комфорта (18 – 20С).
35. Почему основной обмен определяют в стандартных условиях:максимального мышечного и эмоционального покоя, натощак, при температуре комфорта?
Потому что физическая нагрузка, эмоциональное напряжение, прием пищи и изменение температуры окружающей среды увеличивают интенсивность метаболических процессов в организме (расход энергии).
36. На какие процессы расходуется энергия основного обмена в организме?
На обеспечение жизнедеятельности всех органов и тканей организма, клеточный синтез, на поддержание температуры тела.
37. Какие факторы определяют величину должного (среднестатистического) основного обмена здорового человека?
Пол, возраст, рост и масса тела (вес).
38. Какие факторы, кроме пола, веса, роста и возраста, определяют величину истинного (реального) основного обмена здорового человека?
Условия жизнедеятельности, к которым организм адаптирован: постоянное проживание в холодной климатической зоне увеличивает основной обмен; длительное вегетарианское питание – уменьшает.
39. Перечислите способы определения величины должного основного обмена у человека. Какой метод используют для определения величины истинного основного обмена у человека в практической медицине?
По таблицам, по формулам, по номограммам. Метод Крога (неполный газовый анализ).
40. Чему равна величина основного обмена у мужчин и женщин в сутки, а также в расчете на 1 кг массы в сутки?
У мужчин 1500 – 1700 ккал (6300 – 7140 кДж), или 21 – 24 ккал (88 – 101 кДж)/кг/сутки. У женщин примерно на 10% меньше этой величины.
41. Одинакова ли у теплокровных животных и человека величина основного обмена, рассчитанная на 1 м 2 поверхности тела и на 1 кг массы тела?
При расчете на 1м 2 поверхности тела у теплокровных животных разных видов и человека показатели примерно равны, при расчете на 1 кг массы – сильно отличаются.
42. Что называют рабочим обменом?
Совокупность основного обмена и дополнительного расхода энергии, обеспечивающих жизнедеятельность организма в различных условиях.
43. Перечислите факторы, повышающие расход энергии организмом. Что называют специфически-динамическим действием пищи?
Физическая и умственная нагрузка, эмоциональное напряжение, изменение температуры и других условий окружающей среды, специфически-динамическое действие пищи (увеличение расхода энергии после приема пищи).
44. На сколько процентов увеличивается расход энергии организмом после приема белковой и смешанной пищи, жиров и углеводов?
После приема белковой пищи – на 20 – 30%, смешанной пищи – на 10 – 12%.
45. Как влияет температура окружающей среды на расход энергии организмом?
Изменение температуры в интервале 15 – 30С существенно не сказывается на энергозатратах организма. При температуре ниже 15С, а также выше 30С расход энергии увеличивается.
46. Как изменяется обмен веществ при температуре окружающей среды ниже 15? Какое это имеет значение?
Увеличивается. Это предотвращает охлаждение организма.
47. Что называют коэффициентом полезного действия организма при мышечной работе?
Выраженное в процентах отношение энергии, эквивалентной полезной механической работе, ко всей энергии, затраченной на выполнение этой работы.
48. Приведите формулу для расчета коэффициента полезного действия (КПД) у человека при мышечной работе, укажите среднюю его величину, расшифруйте элементы формулы.

где А - энергия, эквивалентная полезной работе, С - общий расход энергии, е - расход энергии за такой же промежуток времени в состоянии покоя. КПД равен 20%.
49. Какие животные называются пойкилотермными и гомойотермными?Пойкилотермные животные (холоднокровные) – с непостоянной температурой тела, зависящей от температуры окружающей среды; гомойотермные (теплокровные) – животные с постоянной температурой тела, не зависящей от температуры окружающей среды.
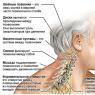
50. Какое значение для организма имеет постоянство температуры тела? В каких органах наиболее интенсивно идет процесс теплообразования?
Обеспечивает высокий уровень жизнедеятельности относительно независимо от температуры окружающей среды. В мышцах, легких, печени, почках.
51. Назовите виды терморегуляции. Сформулируйте суть каждого из них.
Химическая терморегуляция – регуляция температуры тела с помощью изменения интенсивности теплопродукции; физическая терморегуляция – с помощью изменения интенсивности теплоотдачи.
52. Какие процессы обеспечивают теплоотдачу?
Теплоизлучение (радиация), теплоиспарение, теплопроведение, конвекция.
53. Как изменяется просвет сосудов кожи при понижении и при повышении температуры окружающей среды? В чем биологическое значение этого явления?
При понижении температуры сосуды кожи суживаются. При повышении температуры окружающей среды сосуды кожи расширяются. В том, что изменение ширины просвета сосудов, регулируя теплоотдачу, способствует поддержанию постоянной температуры тела.
54. Как и почему изменится теплопродукция и теплоотдача при сильном возбуждении симпатоадреналовой системы?
Теплопродукция увеличится вследствие стимуляции окислительных процессов, а теплоотдача уменьшится в результате сужения кожных сосудов.
55.Перечислите области локализации терморецепторов.
Кожа, кожные и подкожные сосуды, внутренние органы, ЦНС.
56. В каких отделах и структурах ЦНС находятся терморецепторы?
В гипоталамусе, ретикулярной формации среднего мозга, в спинном мозге.
57. В каких отделах ЦНС расположены центры терморегуляции? Какая структура ЦНС является высшим центром терморегуляции?
В гипоталамусе и спинном мозге. Гипоталамус.
58. Какие изменения возникнут в организме при длительном отсутствии в пищевом рационе жиров и углеводов, но при оптимальном поступлении белка с пищей (80 – 100 г в сутки)? Почему?
Будет наблюдаться превышение расхода азота организмом над приходом, потеря веса, поскольку энергозатраты будут покрываться в основном за счет белков и запасов жира, которые не пополняются.
59. В каком количестве и в каком соотношении должны содержаться белки, жиры и углеводы в пищевом рационе взрослого человека (усредненный вариант)?
Белки – 90 г, жиры – 110 г, углеводы – 410 г. Соотношение 1: 1, 2: 4, 6.
60. Как изменяется состояние организма при избыточном поступлении жиров?
Развивается ожирение, атеросклероз (преждевременно). Ожирение является фактором риска развития сердечно-сосудистых заболеваний и их осложнений (инфаркт миокарда, инсульт и др.), снижения продолжительности жизни.
1. Каково соотношение величин основного обмена у детей первых 3 – 4 лет жизни, в период полового созревания, в возрасте 18 – 20 лет и взрослых (ккал/кг/сутки)?
До 3 – 4 лет у детей примерно в 2 раза больше, в период полового созревания – в 1, 5 раза больше, чем у взрослых. В 18 – 20 лет – соответствует норме взрослых.
2. Нарисуйте график изменения основного обмена у мальчиков с возрастом (у девочек основной обмен на 5% ниже).

3. Чем объясняется высокая интенсивность окислительных процессов у ребенка?
Более высоким уровнем метаболизма молодых тканей, относительно большой поверхностью тела и, естественно, большими затратами энергии для поддержания постоянства температуры тела, повышенной секрецией гормонов щитовидной железы и норадреналина.
4. Как изменяются энергетические затраты на рост в зависимости от возраста ребенка: до 3-х месяцев жизни, до начала полового созревания, в период полового созревания?
Увеличиваются в первые 3 месяца после рождения, затем постепенно уменьшаются, а в период полового созревания вновь нарастают.
5. Из чего складывается и как распределяется в процентах общий расход энергии у ребенка в возрасте 1 года по сравнению со взрослым человеком?
У ребенка: 70% приходится на основной обмен, 20% – на движения и поддержание мышечного тонуса, 10% на специфически-динамическое действие пищи. У взрослого: 50 – 40 – 10% соответственно.
6. Взрослые или дети 3 – 5-летнего возраста затрачивают больше энергии при выполнении мышечной работы для достижения одного и того же полезного результата, во сколько раз и почему?
Дети, в 3 – 5 раз, так как у них менее совершенна координация, что приводит к избыточным движениям, в результате чего полезная работа у детей значительно меньше.
7. Как изменяется расход энергии при крике ребенка, на сколько процентов, вследствие чего?
Увеличивается на 100 – 200% вследствие увеличения теплопродукции в результате эмоционального возбуждения и увеличения мышечной активности.
8. Какая часть (в процентах) энерготрат ребенка грудного возраста обеспечивается за счет белков, жиров, углеводов? (сравните с нормой взрослого).
За счет белков – 10%, за счет жиров – 50%, за счет углеводов – 40%. У взрослых – 20 – 30 – 50%, соответственно.
9. Почему дети, особенно в грудном возрасте, быстро перегреваются при повышении температуры окружающей среды? Повышение или понижение температуры окружающей среды дети переносят легче?
Потому, что у детей повышена теплопродукция, недостаточно потоотделение и, следовательно, теплоиспарение, незрелый центр терморегуляции. Понижение.
10. Назовите непосредственную причину и объясните механизм быстрого охлаждения детей (особенно грудного возраста) при понижении температуры окружающей среды.
Повышенная теплоотдача у детей вследствие относительно большой поверхности тела, обильного кровоснабжения кожи, недостаточной теплоизоляции (тонкая кожа, отсутствие подкожной жировой клетчатки) и незрелости центра терморегуляции; недостаточное сужение сосудов.
11. В каком возрасте у ребенка появляются суточные колебания температуры, чем они отличаются от таковых у взрослых, в каком возрасте они достигают нормы взрослого?
В конце 1 месяца жизни; они незначительны и достигают нормы взрослого человека к пяти годам.
12. Что такое температурная "зона комфорта" ребенка, в пределах какой температуры она находится, чему равен этот показатель у взрослых?
Температура внешней среды, при которой индивидуальные колебания температуры кожи ребенка наименее выражены, находится в пределах 21 – 22 о С, у взрослого – 18 – 20 о С.
13. Какие механизмы терморегуляции наиболее готовы к функционированию к моменту рождения? В каких условиях могут включаться механизмы дрожательного термогенеза у новорожденных детей?
Усиленное теплообразование преимущественно недрожательного происхождения (высокий обмен веществ), потоотделение. В условиях предельного холодового воздействия.
14. В каком соотношении должны содержаться белки, жиры и углеводы в пищевом рационе детей в возрасте трех и шести месяцев, 1 года, старше одного года и у взрослых?
До 3 месяцев – 1: 3: 6; в 6 месяцев – 1: 2: 4. В возрасте 1 год и старше – 1: 1, 2: 4, 6, т. е. как и у взрослых.
15. Назовите особенности обмена минеральных солей у детей. С чем это связано?
Наблюдается ретенция солей в организме, особенно повышена потребность в кальции, фосфоре и железе, что связано с ростом организма.
11 Обмен энергии
Непременным условием поддержания жизни является получение организмами энергии из внешней среды, и хотя первоисточник энергии для всего живого - Солнце, непосредственно использовать его излучение способны только растения. Посредством фотосинтеза они превращают энергию солнечных лучей в энергию химических связей. Животные и человек получают необходимую им энергию, поедая растительную пищу. (Для хищных и отчасти для всеядных источником энергии служат другие животные - растительноядные.)
Прямое получение энергии солнечных лучей животными тоже возможно, например, пойкилотермные таким образом поддерживают температуру своего тела. Однако тепло (получаемое из внешней среды и образующееся в самом организме) не может быть преобразовано ни в какой другой вид энергии. Живые организмы, в отличие от технических устройств, принципиально неспособны к этому. Машина, использующая энергию химических связей, (например, двигатель внутреннего сгорания), сначала превращает ее в тепло и только затем - в работу: химическая энергия топлива → тепло → работа (расширение газа в цилиндре и движение поршня). В живых организмах возможна только такая схема: химическая энергия → работа.
Итак, энергия химических связей в молекулах пищевых веществ - практически единственный источник энергии для животного организма, а тепловая энергия может быть использована им только для поддержания температуры своего тела. Кроме того, тепло из-за быстрого рассеивания в окружающей среде не может быть и запасено в организме на длительный срок. Если возникает избыток тепла в теле, то для гомойотермных животных это становится серьезной проблемой и иногда даже угрожает их жизни (см. разд. 11.3).
11.1. Источники энергии и пути ее превращения в организме
Живой организм - открытая энергетическая система: он получает из окружающей среды энергию (почти исключительно в виде химических связей), преобразует ее в тепло или работу и в таком виде возвращает ее в окружающую среду.
Компоненты пищевых веществ, поступающие из желудочно-кишечного тракта в кровь (например, глюкоза, жирные кислоты или аминокислоты), сами по себе не способны непосредственно передавать энергию своих химических связей ее потребителям, например, калий-натриевому насосу или актину и миозину мышц. Между пищевыми «энергоносителями» и «потребителями» энергии есть универсальный посредник - аденозинтрифосфат (АТФ). Именно он является непосредственным источником энергии для любых процессов в живом
организме. Молекула АТФ представляет собой соединение аденина, рибозы и трех фосфатных групп (рис. 11.1).
Связи между кислотными остатками (фосфатами) содержат в себе значительное количество энергии. Отщепляя под действием фермента АТФазы концевой фосфат, АТФ превращается в аденозиндифосфат (АДФ). При этом высвобождается 7,3 ккал/моль энергии. Энергия химических связей в молекулах пищевых веществ используется для ресинтеза АТФ из АДФ. Рассмотрим этот процесс на примере глюкозы (рис. 11.2).
Первый этап утилизации глюкозы - гликолиз. В ходе его молекула глюкозы сначала превращается в пировиноградную кислоту (пиру ват), давая при этом энергию для ресинтеза АТФ. Затем пируват превращается в ацетилкоэнзим А - исходный продукт для следующего этапа утилизации - цикла Кребса. Многократные превращения веществ, составляющие суть этого цикла, дают дополнительную энергию для ресинтеза АТФ и заканчиваются высвобождением ионов водорода. С передачи этих ионов в дыхательную цепь начинается третий этап - окислительное фосфорилирование, в результате которого также образуется АТФ.
В совокупности все три этапа утилизации (гликолиз, цикл Кребса и окислительное фосфорилирование) составляют процесс тканевого дыхания. Принципиально важно, что первый этап (гликолиз) проходит без использования кислорода (анаэробное дыхание) и приводит к образованию лишь двух молекул АТФ. Два последующих этапа (цикл Кребса и окислительное фосфорилирование) могут происходить только в кислородной среде (аэробное дыхание). Полная утилизация одной молекулы глюкозы приводит к появлению 38 молекул АТФ.
Существуют организмы, не только не нуждающиеся в кислороде, но погибающие в кислородной (или воздушной) среде - облигатные анаэробы. К ним, например, относятся бактерии - возбудители газовой гангрены (Clostridium perfringes), столбняка (С. tetani), ботулизма (С. botulinum) и др.
У животных анаэробные процессы являются вспомогательным видом дыхания. Например, при интенсивных и частых сокращениях мышц (или при статическом их сокращении) доставка кислорода кровью отстает от потребностей мышечных клеток. В это время образование АТФ происходит анаэробным путем с накоплением пирувата, который превращается в молочную кислоту (лактат). Нарастает кислородный долг. Прекращение или ослабление мышечной работы устраняет несоответствие между потребностью ткани в кислороде и возможностями его доставки, лактат превращается в пируват, последний либо через стадию ацетилкоэнзима А окисляется в цикле Кребса до двуокиси углерода, либо путем глюконеогенеза переходит в глюкозу.
Согласно второму началу термодинамики всякое превращение энергии из одного вида в другой происходит с обязательным образованием значительного количества тепла, которое затем рассеивается в окружающем пространстве. Поэтому синтез АТФ и передача энергии от АТФ к собственно «потребителям энергии» происходят с потерей примерно половины ее в виде тепла. Упрощая, можно представить эти процессы следующим образом (рис. 11.3).
Примерно половина химической энергии, содержащейся в пище, сразу же превращается в тепло и рассеивается в пространстве, другая половина идет на образование АТФ. При последующем расщеплении АТФ половина высвободившейся энергии опять-таки превращается в тепло. В результате на выполнение внешней работы (например, бег или перемещение каких-либо предметов в пространстве) животное и человек могут затратить не более 1/4 всей потребленной в виде пищи энергии. Таким образом, коэффициент полезного действия высших животных и человека (около 25%) в несколько раз выше, чем, например, коэффициент полезного действия (КПД) паровой машины.
Вся внутренняя работа (кроме процессов роста и накопления жира) быстро превращается в тепло. Примеры: (а) энергия, вырабатываемая сердцем, превращается в тепло благодаря сопротивлению сосудов току крови; (б) желудок выполняет работу по секреции соляной кислоты, поджелудочная железа секретирует гидрокарбонат-ионы, в тонкой кишке эти вещества взаимодействуют, и заложенная в них энергия преобразуется в тепло.
Результаты внешней (полезной) работы, произведенной животным или человеком, также в конечном счете превращаются в тепло: перемещение тел в пространстве согревает воздух, возведенные сооружения рушатся, отдавая заложенную в них энергию земле и воздуху в виде тепла. Египетские пирамиды - редкий пример того, как энергия мышечного сокращения, затраченная почти 5000 лет назад, все еще ждет неизбежного превращения в тепло.
Уравнение энергетического баланса:
Е = А + Н + S,
где Е - общее количество энергии, получаемой организмом с пищей; А - внешняя (полезная) работа; Н - теплоотдача; S - запасенная энергия.
Потери энергии с мочой, кожным салом и др. выделениями крайне малы, и ими можно пренебречь.
Дыхательный коэффициент составляет 18,10:24,70 = 0,73.[ ...]
Дыхательный коэффициент при нормальном созревании плодов не остается постоянным. В предклимакте-рической стадии он равен примерно 1 и по мере созревания достигает значений 1,2... 1,5. При отклонениях на ±0,25 от единицы в плодах еще не наблюдаются аномалии в обмене веществ, и только при больших отклонениях можно предполагать физиологические расстройства. Интенсивность дыхания отдельных слоев тканей какого-либо плода неодинакова. В соответствии с большей активностью ферментов в кожице в ней отмечается во много раз большая интенсивность дыхания, чем у паренхимной ткани (Хулме и Родс, 1939). При снижении содержания кислорода и увеличении концентрации двуокиси углерода в паренхимных клетках по мере удаления от кожицы к сердцевине плода интенсивность дыхания падает.[ ...]
Прибор для определения дыхательного коэффициента, пинцеты, полоски фильтровальной бумаги, песочные часы па 2 мин, стеклянные чашки, пипетки, стеклянные палочки, конические колбы на 250 мл.[ ...]
Прибор для определения дыхательного коэффициента состоит из большой пробирки с плотно пригнанной каучуковой пробкой, в которую вставлена изогнутая под прямым углом измерительная трубка со шкалой из миллиметровой бумаги.[ ...]
Потребление кислорода и коэффициент его утилизации были постоянными при снижении р02 до 60 и 20% исходного (в зависимости от скорости тока). При концентрации кислорода, слегка превышающей критическую, длительное время поддерживался максимальный объем вентиляции (в течение нескольких часов). Объем вентиляции возрастал при этом в 5,5 раза, но в отличие от карпов он снижался начиная с 22% уровня насыщения воды кислородом. Авторы считают , что снижение объема вентиляции у рыб при экстремальной гипоксии - следствие кислородной недостаточности дыхательной мускулатуры. Соотношения частоты дыхания и сердцебиений были равны 1,4 в норме и 4,2 при дефиците кислорода.[ ...]
Вводные пояснения. Преимущества метода: высокая чувствительность, что позволяет работать с малыми навесками опытного материала; возможность наблюдать за динамикой газообмена и одновременно учитывать газообмен 02 и С02, что позволяет установить дыхательный коэффициент.[ ...]
Поэтому значение pH в окситеике снижается почти до 6,0, в то время как в аэротенке рН>7Д При максимальной нагрузке потребляемая мощность для окситенка, включающая мощность оборудования для получения кислорода 1,3 м3/ (л.с.-ч) и мощность аэратора (рис. 26.9), должна быть меньше, чем мощность аэратора для аэротенка. Это объясняется высокой концентрацией кислорода (выше 60%) ка всех ступенях окситенка.[ ...]
Динамика выделения углекислого газа (С?со2), поглощения кислорода ([ ...]
Морские и пресноводные рыбы в данных условиях опыта имели примерно одинаковый дыхательный коэффициент (ДК). Недостатком этих данных является то, что автор взял для сравнения золотую рыбку, которая вообще мало потребляет кислорода и вряд ли может служить эталоном сравнения.[ ...]
В отношении газообмена насекомых, находящихся в зимней спячке, следует сказать, что и дыхательный коэффициент при этом понижается1. Так, например, Драйер (1932) обнаружил, что у муравья Formica ulkei Emery в активном состоянии дыхательный коэффициент был равен 0,874; когда же муравьи перед зимней спячкой стали малоподвижными, дыхательный коэффициент снизился до 0,782, и в период спячки снижение достигло 0,509-0,504. У колорадского картофельного жука Leptinotarsa decemlineata Say. в период зимовки дыхательный коэффициент снижается до 0,492-0,596, тогда как в летнее время он равен 0,819- 0,822 (Ушатинская, 1957). Это объясняется тем, что в активном состоянии насекомые живут преимущественно за счет белковой и углеводной пищи, в спячке же расходуется главным образом жир, требующий для окисления меньшего количества кислорода.[ ...]
В герметичных резервуарах, рассчитанных на давление в ГП рк. д = 1962 Па (200 мм. вод. ст), при высоких коэффициентах оборачиваемости продолжительность простаивания резервуара с «мертвым» остатком до начала заполнения может быть так мала, что дыхательный клапан не успеет открыться для «выдоха». Тогда потери от «обратного.выдоха» отсутствуют.[ ...]
Для понимания биохимических процессов, протекающих в организме, большое значение имеет величина дыхательного коэффициента. Дыхательный коэффициент (ДК) - отношение выдыхаемой угольной кислоты к потребляемому кислороду.[ ...]
Для суждения о влиянии температуры на какой-либо процесс обычно оперируют величиной температурного коэффициента. Температурный коэффициент (£>ю) процесса дыхания зависит от типа растений и от градаций температуры. Так, при повышении температуры от 5 до 15°С 0 ю может возрастать до 3, тогда как повышение температуры от 30 до 40°С увеличивает интенсивность дыхания менее значительно (Фю около 1,5). Большое значение имеет фаза развития растений. По данным Б, А. Рубина, на каждой фазе развития растений для процесса дыхания наиболее благоприятны те температуры, на фоне которых обычно проходит эта фаза, Изменение оптимальных температур при дыхании растений в зависимости от фазы их развития связано с тем, что в процессе онтогенеза меняются пути дыхательного обмена. Между тем для разных ферментных систем наиболее благоприятным являются разные температуры. В этой связи интересно, что в более поздние фазы развития растений наблюдаются случаи, когда фла-виновые дегидрогеназы выступают в роли конечных оксидаз, передавая водород непосредственно кислороду воздуха.[ ...]
Все исследованные рыбы в неволе потребляют кислорода меньше, чем в природных условиях. Слабое увеличение дыхательного коэффициента у рыб, содержавшихся в аквариумах, свидетельствует об изменении качественной стороны обмена в сторону большего участия в нем углеводов и белков. Автор объясняет это худшим кислородным режимом аквариума по сравнению с естественными условиями; кроме того, в аквариуме рыбы малоподвижны.[ ...]
Для снижения выброса паров вредных веществ применяются также диски-отражатели, устанавливаемые под монтажным патрубком дыхательного клапана. При высоком коэффициенте оборачиваемости атмосферных резервуаров эффективность дисков-отражателей может достичь 20-30%.[ ...]
Донасыщение ГП может происходить после заполнения, если газовое пространство было не вполне насыщено парами. В этом случае дыхательный клапан после заполнения емкости не закрывается и сразу начинается дополнительный выдох. Такое явление встречается в резервуарах, имеющих высокий коэффициент оборачиваемости или заполняемых частично, не до предельной высоты взлива, а также в резервуарах с замедленными процессами насыщения ГП (резервуары с понтонами и заглубленные). Донасыщение ГП особенно характерно для резервуаров, впервые заполненных после зачистки и проветривания. Такой вид потерь иногда называют потерями от насыщения или донасыщения ГП.[ ...]
При известных и и0 Аcjcs можно также определять по графикам, аналогичным приведенному на рис. 14. В методиках расчета потерь приведены подобные графики для типовых резервуаров РВС, различных типов дыхательных клапанов и их количества. Величина Аc/cs означает прирост концентрации в ГП за суммарное время простоя (тп) и заполнения резервуара (тэ), т. е. т = т„ + т3; ее определяют приближенно по графикам (см. рис. 3). При использовании формулы (!9) необходимо иметь б виду, что при полном насыщении ГП ccp/cs = 1 и что время полного насыщения ГП наземных резервуаров ограничено 2-4 сутками (в зависимости от состояния погоды и других условий), а график на" рис. 3 приближенный. Поэтому, получив по формуле (19) значения ccp/cs>l, что означает наступление полного насыщения ГП ранее момента конца простоя или конца заполнения резервуара, в формулу (18) необходимо подставлять ccp/cs = 1.[ ...]
Оценим количественные связи между этими двумя потоками газов. Во-первых» отношение объема выделенного углекислого газа к объему потребленного кислорода (дыхательный коэффициент) для большинства сточных вод и активных илов меньше единицы. Во-вторых, объемные коэффициенты массопередачи для кислорода и углекислого газа близка друг к другу. В-третьих, константа фазового равновесия углекислого газа почти в 30 раз меньше кислородной. В-четвертых, углекислый газ не только присутствует в иловой смеси в растворенном состояний, но и вступает с водой в химическое взаимодействие.[ ...]
При сравнении обоих типов дыхания бросается в глаза неодинаковое отношение поглощения кислорода к выделению двуокиси углерода. Отношение СО2/О2 обозначается как дыхательный коэффициент КО.[ ...]
Если в процессе дыхания окисляются органические вещества с относительно более высоким содержанием кислорода, чем в углеводах, например органические кислоты - щавелевая, винная и их соли, то дыхательный коэффициент будет значительно больше 1. Он также будет больше 1 в том случае, когда часть кислорода, используемого для дыхания микробов, берется из углеводов; или же при дыхании тех дрожжей, у которых одновременно с аэробным дыханием происходит спиртовое брожение. Если же наряду с аэробным дыханием протекают другие процессы, при которых используется добавочный кислород, то дыхательный коэффициент будет меньше 1. Он будет меньше 1 и тогда, когда в процессе дыхания окисляются вещества с относительно небольшим содержанием кислорода, например белки, углеводороды и др. Следовательно, зная значение дыхательного коэффициента, можно определить, какие вещества окисляются в процессе дыхания.[ ...]
Наиболее общий показатель скорости окисления- интенсивность дыхания, о которой можно судить по поглощению кислорода, выделению диоксида углерода и окислению органического вещества. Другие показатели дыхательного метаболизма: величина дыхательного коэффициента, соотношение гликолитического и пенто-зофосфатного путей распада сахаров, активность окислительно-восстановительных ферментов. Об энергетической эффективности дыхания можно судить по интенсивности окислительного фосфорилирования митохондрий.[ ...]
Показанные на яблоках сорта Кокс оранж тенденции в отношении влияния концентраций кислорода и двуокиси углерода в воздухе камеры действительны для всех других сортов яблок, за исключением случаев более сильного увеличения дыхательного коэффициента при понижении температуры.[ ...]
Величина ДК зависит и от других причин. В некоторых тканях из-за затрудненного доступа кислорода наряду с аэробным происходит анаэробное дыхание, не сопровождающееся поглощением кислорода, что приводит к повышению значения ДК. Величина коэффициента обусловлена также полнотой окисления дыхательного субстрата. Если, кроме конечных продуктов, в тканях накапливаются менее окисленные соединения (органические кислоты), то ДК[ ...]
Количественные определения зависимости газообмена у рыб от температуры были проведены многими исследователями. В большинстве случаев изучение этого вопроса ограничивалось преимущественно количественной стороной дыхания - величиной дыхательного ритма, величиной потребления кислорода и затем вычислением температурных коэффициентов при разных температурах.[ ...]
Для снижения потерь в результате испарения и загрязнения воздушной среды, резервуары для бензина оснащают газовой обвязкой, соединяющей воздушные пространства резервуаров, в которых хранится продукция одной марки, а также устанавливают общий дыхательный клапан. Описанные выше «большие и малые дыхания», вентиляция газового пространства, также являются причиной загрязнения воздушной среды при хранении нефтепродуктов на сельскохозяйственных объектах, так как при коэффициенте оборачиваемости резервуарного парка 4-6 коэффициент оборачиваемости запасов топлива составляет 10-20, а это означает снижение коэффициента использования резервуаров 0,4-0,6. В целях предотвращения загрязнения атмосферы на нефтескладах предусматривают очистные устройства и бензомаслоуловители.[ ...]
Полученные к настоящему времени данные показывают, что экстремальные температуры вызывают угнетение деятельности физиологической системы, в частности транспорта газов у рыб. При этом развивается бра-дикардия, нарастает аритмия, снижаются потребление кислорода и коэффициент его утилизации. Вслед за этими изменениями работы кардиореспира-торного аппарата постепенно прекращается вентиляция жабр и в последнюю очередь перестает функционировать миокард. По-видимому, аноксия дыхательной мускулатуры и общая кислородная недостаточность одна из причин гибели рыб при перегреве. Повышение температуры приводит к ускорению утилизации кислорода и, как следствие, к падению его напряжения в дорзальной аорте, что, в свою очередь, служит сигналом для усиления вентиляции жабр .[ ...]
Перед применением модели следует провести проверку ее кинетических параметров. Проверка модели системы с чистым кислородом для очистки бытовых и промышленных сточных вод была сделана Мюллером и др. (1]. При проверке моделей для очистки бытовых сточных вод использовали дыхательный коэффициент Я.С, равный 1,0, тогда как для промышленных сточных вод он составляет 0,85 и даже 0,60. Дополнительная проверка химических взаимодействий была сделана совсем недавно при изучении сточных вод целлюлозно-бумажной фабрики (рис.26.6). Для оценки полученных данных дыхательный коэффициент был принят равным 0,90. Хотя данных о содержании аммонийного азота было не так много, была отмечена бюлее низкая потребность в нем для роста микроорганизмов, чем это традиционно наблюдалось в биологических системах.[ ...]
Для решения вопроса о сущности действия температуры на обмен веществ рыбы нужно знать не только степень увеличения или уменьшения обмена с изменением температуры, но и качественные изменения в отдельных звеньях, составляющих обмен веществ. Качественную сторону обмена до некоторой степени могут характеризовать такие коэффициенты, как дыхательный и аммиачный (отношение выделенного аммиака как конечного продукта азотистого обмена к потребленному кислороду) (рис. 89).[ ...]
Из приведенного выше уравнения (4) следует, что отношение констант для 02 и С02 равно 1,15 , т. е. использование С02-техники измерения баланса, казалось бы, позволяет проводить наблюдения при несколько более высоких значениях 2 и соответственно больших скоростях течения. Но это кажущееся преимущество исчезает, если принять, что дыхательный коэффициент меньше 1. Кроме того, как показал Таллинг 32], точность определения С02 в природных водах не может быть лучше чем ± 1 мкмоль/л (0,044 мг/л), а кислорода - ±0,3 ,мкмоль/л (0,01 мг/л). Следовательно, даже если принять дыхательный коэффициент равным 1, точность балансового метода, основанного на учете баланса кислорода, оказывается по крайней мере втрое выше, чем при определении углекислоты.[ ...]
Морфо-физиологический метод применялся в наших исследованиях с некоторыми дополнениями. Это позволило определять с достаточной точностью (±3,5%) количества поглощенного кислорода, выделенной углекислоты и дыхательный коэффициент (ДК) на целых проростках 10 -12 дневного возраста и листьях растений из полевых опытов. Принцип этой методики заключается в том, что растения помещенные в замкнутый сосуд (газовая пипетка специальной конструкции) с атмосферным воздухом, в результате дыхания изменяют состав воздуха. Таким образом, зная объем сосуда и определив процентный состав воздуха в начале и в конце опыта, нетрудно рассчитать количество поглощенного 02:и выделенной СОг растениями.[ ...]
Различные органы и ткани растения сильно различаются по условиям снабжения их кислородом. В листе кислород свободно поступает практически к каждой клетке. Сочные плоды, корнеплоды, клубни вентилируются очень плохо; они слабо проницаемы для газов, не только для кислорода, но и для углекислого газа. Естественно, в этих органах процесс дыхания сдвигается в анаэробную сторону, дыхательный коэффициент возрастает. Возрастание дыхательного коэффициента и сдвиг процесса дыхания в анаэробную сторону наблюдаются в меристематических тканях. Таким образом, разные органы характеризуются не только различной интенсивностью, но и неодинаковым качеством дыхательного процесса.[ ...]
Вопрос о веществах, используемых в процессе дыхания, издавна еаниман физиологов. Еще в работах И. П. Бородина было показано, что интенсивность процесса дыхания прямо пропорциональна содержанию в тканях растений углеводов. Это дало основание предположить, что именно углеводы яеляются основным веществом, потребляемым при дыхании. В выяснении данного вопроса большое значение имеет определение дыхательного коэффициента. Дыхательный коэффициент - это объемное или молярное отношение СОг, выделившегося в процессе дыхания, к поглощенному за этот же промежуток времени Ог- При нормальном доступе кислорода величина дыхательного коэффициента зависит от субстрата дыхания. Если в процессе дыхания используются углеводы, то процесс идет согласно уравнению СеН) 2О5+6О2=6СО2+6Н2О, в этом случае дыхательный коэффициент равен единице!=1. Однако, если разложению в процессе дыхания подвергаются более окисленные соединения, например органические кислоты, поглощение кислорода уменьшается, дыхательный коэффициент становится больше единицы. При окислении в процессе дыхания более восстановленных соединений, таких, как жиры или белки, требуется больше кислорода и дыхательный коэффициент становится меньше единицы.[ ...]
Итак, простейший процесс аэробного дыхания представляется в следующем виде. Молекулярный кислород, потребляемый в процессе дыхания, используется в основном для связывания водорода, образующегося при окислении субстрата. Водород от субстрата передается к кислороду через ряд промежуточных реакций, проходящих последовательно с участием ферментов и переносчиков. Определенное представление о характере процесса дыхания дает так называемый дыхательный коэффициент. Под этим понимают отношение объема выделившегося углекислого газа к объему кислорода, поглощенного в процессе дыхания (С02:02).[ ...]
Эффективность работы кардиореспираторного аппарата рыб, его резервные возможности, лабильность частотных и амплитудных показателей зависят от видовой принадлежности и экологических особенностей рыб. При повышении температуры на одну и ту же величину (с 5 до 20°С) частота дыхания у судака увеличилась с 25 до 50 в минуту, у щуки с 46 до 75, у язя с 63 до 112 в минуту. Потребление кислорода увеличивается параллельно с увеличением частоты, но не глубины дыхания. Наибольшее число дыхательных движений для прокачивания единицы объема воды производит подвижный язь, а наименьшее - менее активный оксифильный судак, что положительно коррелирует с интенсивностью газообмена у исследованных видов . По мнению авторов, соотношение максимального объема вентиляции и соответствующего ему коэффициента утилизации кислорода определяет максимальные энергетические возможности организма. В покое наибольшие интенсивность газообмена и объем вентиляции были у оксифильного судака, а при функциональной нагрузке (двигательная активность, гипоксия) -- у язя. При низкой температуре прирост объема вентиляции у язя в ответ на гипоксию был больше, чем при высокой, а именно: 20-кратный при 5°С и 8-кратный при 20°С. У ОггИоЬоп тюго1ер1с1оги5 при гипоксии (40% насыщения) объем воды, прокачиваемой через жабры, изменяется в меньшей степени: при 12°С увеличивается в 5 раз, а при 28°С - в 4,3 раза .[ ...]
Значительно менее полно исследованы показатели углеводного обмена при адаптивной экзогенной гипоксии, т. е. при легком и умеренном дефиците кислорода в окружающей среде. Однако имеющиеся немногочисленные экспериментальные данные показывают, что и в этом случае происходят усиленное использование гликогена в мышцах, повышение содержания молочной кислоты и сахара в крови. Как и следовало ожидать, уровень насыщения воды кислородом, при котором отмечаются эти сдвиги, неодинаков для разных видов. Например, у миноги отмечена гипергликемия при снижении содержания кислорода всего лишь на 20% от исходного и у 1 аЬео сарепвк концентрация сахара в крови оставалась постоянно низкой даже при 40% насыщения воды кислородом и только дальнейшее снижение насыщения приводило к быстрому увеличению уровня сахара в крови. Повышение сахара в крови и молочной кислоты отмечено при гипоксии у линя . Аналогичная реакция на гипоксию отмечена и у канального сомика . В первой из этих работ при 50%-ном насыщении воды кислородом у рыб выявлено повышение содержания молочной кислоты, которое продолжалось и в первый час нормоксии, т. е. после возвращения рыб в нормальные кислородные условия. Восстановление биохимических показателей к норме происходило в течение 2-6 ч, а увеличение содержания лактата и дыхательного коэффициента с 0,8 до 2,0 свидетельствовало об увеличении анаэробного гликолиза.
Дыхательный коэффициент (ДК) представляет собой отношение объема выделенного углекислого газа к объему поглощенного кислорода за определенное время. Если в процессе обмена в организме окисляются только углеводы, то дыхательный коэффициент будет равен 1. Это видно из следующей формулы:
Следовательно, для образования одной молекулы СО 2 при обмене углеводов потребуется одна молекула О 2 . Так как по закону Авогадро-Жерара равные количества молекул при одинаковой температуре и давлении занимают равные объемы. Следовательно, дыхательный коэффициент при окислении углеводов будет равен 1:
![]()
Для жиров он будет:
![]()
На окисление одной молекулы жира необходимо 81,5 молекулы кислорода, а на окисление 1 грамм-молекулы жира нужно 81,5 х 22,4 л кислорода, то есть 1825,6 л О 2 , где 22,4 – объем одной грамм-молекулы в литрах. Грамм-молекула жира равна 890 г, тогда 1 л кислорода окисляет ![]() 487 г жира. 1 г жира при полном окислении выделяет 38,945 кДж (9,3 ккал)*, а 0,487 дает 18,551 кДж. Следовательно, калорический эквивалент 1 л кислорода при дыхательном коэффициенте 0,7 будет равен 18,551 кДж. В нормальных условиях дыхательный коэффициент лежит в пределах 1 и 0,7. При ДК 0,7 в организме окисляются жиры и при этом калорический эквивалент, или тепловая ценность 1 л кислорода, равен 18,551 кДж, а при ДК=1 – 21,135.
487 г жира. 1 г жира при полном окислении выделяет 38,945 кДж (9,3 ккал)*, а 0,487 дает 18,551 кДж. Следовательно, калорический эквивалент 1 л кислорода при дыхательном коэффициенте 0,7 будет равен 18,551 кДж. В нормальных условиях дыхательный коэффициент лежит в пределах 1 и 0,7. При ДК 0,7 в организме окисляются жиры и при этом калорический эквивалент, или тепловая ценность 1 л кислорода, равен 18,551 кДж, а при ДК=1 – 21,135.