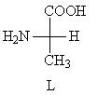
Митохондрии биологическая роль. Строение и функции митохондрий
Велика. По своему строению они представляют собой обычно сферические органеллы, встречающиеся в эукариотической клетке в количестве от нескольких сот до 1-2 тысяч и занимающие 10-20 % её внутреннего объёма. Сильно варьируют так же размеры (от 1 до 70 мкм) и форма митохондрий. В зависимости от того, в каких участках клетки в каждый конкретный момент происходит повышенное потребление энергии, митохондрии способны перемещаться по цитоплазме в зоны наибольшего энергопотребления, используя для движения структуры цитоскелета эукариотической клетки. В клетках растений и животных одновременно существуют примерно в равных количествах три типа митохондриальных органелл: молодые протомитохондрии, зрелые митохондрии и старые постмитохондрии, деградирующие в липофусциновые гранулы .
Структура митохондрий
Наружная мембрана
Наружная мембрана митохондрии имеет толщину около 7 нм, не образует впячиваний и складок, и замкнута сама на себя. На наружную мембрану приходится около 7 % от площади поверхности всех мембран клеточных органелл. Основная функция - отграничение митохондрии от цитоплазмы . Наружная мембрана митохондрии состоит из липидов с вкраплениями белков (соотношение 2: 1). Особую роль играет порин - каналообразующий белок : он формирует в наружной мембране отверстия диаметром 2-3 нм, через которые могут проникать небольшие молекулы и ионы весом до 5 кДа . Крупные молекулы могут пересекать наружную мембрану только посредством активного транспорта через транспортные белки митохондриальных мембран. Для наружной мембраны характерно присутствие ферментов : монооксигеназы, ацил-СоА-синтетазы и фосфолипазы А 2 . Наружная мембрана митохондрии может взаимодействовать с мембраной эндоплазматического ретикулума ; это играет важную роль в транспортировке липидов и ионов кальция .
Межмембранное пространство
Межмембранное пространство представляет собой пространство между наружной и внутренней мембранами митохондрии. Его толщина - 10-20 нм. Так как наружная мембрана митохондрии проницаема для небольших молекул и ионов, их концентрация в периплазматическом пространстве мало отличается от таковой в цитоплазме. Напротив, крупным белкам для транспорта из цитоплазмы в периплазматическое пространство необходимо иметь специфические сигнальные пептиды ; поэтому белковые компоненты периплазматического пространства и цитоплазмы различны. Одним из белков, содержащихся не только во внутренней мембране, но и в периплазматическом пространстве, является цитохром c .
Внутренняя мембрана
Энергетический потенциал (запас энергии) в молекуле убихинола существенно ниже, чем в молекуле НАДН, а разница в подобной энергии временно запасается в виде электрохимического протонного градиента. Последний возникает в результате того, что перенос электронов по простетическим группам комплекса I, ведущий к снижению энергетического потенциала электронов, сопровождается трансмембранным переносом двух протонов из матрикса в межмембранное пространство митохондрии.
Восстановленный убихинол мигрирует в плоскости мембраны, где достигает второго фермента дыхательной цепи - комплекса III (цитохром bc 1 ). Последний представляет собой димер с молекулярной массой более 300 кДа, сформированный из восьми полипептидных цепей и содержащий атомы железа как в виде железосерных центров, так и в виде комплексов с гемами b (I), b (II) и c 1 - сложными гетероциклическими молекулами с четырьмя атомами азота , расположенными по углам металлосвязывающего квадрата. Комплекс III катализирует реакцию окисления двух убихинолов до убихинон ов, восстанавливая две молекулы цитохрома c (гемсодержащий переносчик, находящегося в межмембранном пространстве). Отщепляющиеся при этом от убихинолов четыре протона освобождаются в межмембранное пространство, продолжая формирование электрохимического градиента.
Последняя стадия катализируется комплексом IV (цитохром c -оксидаза) с молекулярной массой около 200 кДа, состоящим из 10-13 полипептидных цепей и, помимо двух различных гемов, включающим также несколько атомов меди , прочно связанных с белками. При этом электроны, отбираемые у восстановленного цитохрома c , пройдя по атомам железа и меди в составе комплекса IV, попадают на связанный в активном центре этого фермента кислород, что приводит к образованию воды .
Таким образом, суммарная реакция, катализируемая ферментами дыхательной цепи, состоит в окислении НАДН кислородом с образованием воды. По сути этот процесс заключается в ступенчатом переносе электронов между атомами металлов , присутствующих в простетических группах белковых комплексов дыхательной цепи, где каждый последующий комплекс обладает более высоким сродством к электрону, чем предыдущий. При этом сами электроны передаются по цепи до тех пор, пока не соединятся с молекулярным кислородом, обладающим наибольшим сродством к электронам. Освобождаемая же при этом энергия запасается в виде электрохимического (протонного) градиента по обе стороны внутренней мембраны митохондрий. При этом считается, что в процессе транспорта по дыхательной цепи пары электронов перекачивается от трёх до шести протонов.
Завершающим этапом функционирования митохондрии является генерация АТФ , осуществляемая встроенным во внутреннюю мембрану специальным макромолекулярным комплексом с молекулярной массой 500 кДа. Этот комплекс, называемый АТФ-синтазой, как раз и катализирует синтез АТФ путём конверсии энергии трансмембранного электрохимического градиента протонов водорода в энергию макроэргической связи молекулы АТФ.
АТФ-синтаза
В структурно-функциональном плане АТФ-синтаза состоит из двух крупных фрагментов, обозначаемых символами F 1 и F 0 . Первый из них (фактор сопряжения F 1) обращён в сторону матрикса митохондрии и заметно выступает из мембраны в виде сферического образования высотой 8 нм и шириной 10 нм. Он состоит из девяти субъединиц, представленных пятью типами белков. Полипептидные цепи трёх субъединиц α и стольких же субъединиц β уложены в похожие по строению белковые глобулы , которые вместе образуют гексамер (αβ) 3 , имеющий вид слегка приплюснутого шара. Подобно плотно уложенным долькам апельсина, последовательно расположенные субъединицы α и β образуют структуру, характеризующуюся осью симметрии третьего порядка с углом поворота 120°. В центре этого гексамера находится субъединица γ, которая образована двумя протяжёнными полипептидными цепями и напоминает слегка деформированный изогнутый стержень длиной около 9 нм. При этом нижняя часть субъединицы γ выступает из шара на 3 нм в сторону мембранного комплекса F 0 . Также внутри гексамера находится минорная субъединица ε, связанная с γ. Последняя (девятая) субъединица обозначается символом δ и расположена на внешней стороне F 1 .
Мембранная часть АТФ-синтазы, называемая фактором сопряжения F 0 , представляет собой гидрофобный белковый комплекс, пронизывающий мембрану насквозь и имеющий внутри себя два полуканала для прохождения протонов водорода. Всего в состав комплекса F 0 входит одна белковая субъединица типа а , две копии субъединицы b , а также от 9 до 12 копий мелкой субъединицы c . Субъединица а (молекулярная масса 20 кДа) полностью погружена в мембрану, где образует шесть пересекающих её α-спиральных участков. Субъединица b (молекулярная масса 30 кДа) содержит лишь один сравнительно короткий погружённый в мембрану α-спиральный участок, а остальная её часть заметно выступает из мембраны в сторону F 1 и закрепляется за расположенную на её поверхности субъединицу δ. Каждая из 9-12 копий субъединицы c (молекулярная масса 6-11 кДа) представляет собой сравнительно небольшой белок из двух гидрофобных α-спиралей, соединённых друг с другом короткой гидрофильной петлёй, ориентированной в сторону F 1 , а все вместе образуют единый ансамбль, имеющий форму погружённого в мембрану цилиндра. Выступающая из комплекса F 1 в сторону F 0 субъединица γ как раз и погружена внутрь этого цилиндра и достаточно прочно зацеплена за него.
Таким образом, в молекуле АТФ-синтазы можно выделить две группы белковых субъединиц, которые могут быть уподоблены двум деталям мотора: ротору и статору . «Статор» неподвижен относительно мембраны и включает в себя шарообразный гексамер (αβ) 3 , находящуюся на его поверхности и субъединицу δ, а также субъединицы a и b мембранного комплекса F 0 . Подвижный относительно этой конструкции «ротор» состоит из субъединиц γ и ε, которые, заметно выступая из комплекса (αβ) 3 , соединяются с погружённым в мембрану кольцом из субъединиц c .
Способность синтезировать АТФ - свойство единого комплекса F 0 F 1 , сопряжённого с переносом протонов водорода через F 0 к F 1 , в последнем из которых как раз и расположены каталитические центры, осуществляющие преобразование АДФ и фосфата в молекулу АТФ. Движущей же силой для работы АТФ-синтазы является протонный потенциал, создаваемый на внутренней мембране митохондрий в результате работы цепи электронного транспорта.
Сила, приводящая в движение «ротор» АТФ-синтазы, возникает при достижении разности потенциалов между наружной и внутренней сторонами мембраны > 220 мВ и обеспечивается потоком протонов, протекающих через специальный канал в F 0 , расположенный на границе между субъединицами a и c . При этом путь переноса протонов включает в себя следующие структурные элементы:
- Два расположенных несоосно «полуканала», первый из которых обеспечивает поступление протонов из межмембранного пространства к существенно важным функциональным группам F 0 , а другой обеспечивает их выход в матрикс митохондрии;
- Кольцо из субъединиц c , каждая из которых в своей центральной части содержит протонируемую карбоксильную группу, способную присоединять H + из межмембранного пространства и отдавать их через соответствующие протонные каналы. В результате периодических смещений субъединиц с , обусловленных потоком протонов через протонный канал происходит поворот субъединицы γ, погружённой в кольцо из субъединиц с .
Таким образом, каталитическая активность АТФ-синтазы непосредственно связана с вращением её «ротора», при котором поворот субъединицы γ вызывает одновременное изменение конформации всех трёх каталитических субъединиц β, что в конечном счёте и обеспечивает работу фермента. При этом в случае образования АТФ «ротор» крутится по часовой стрелке со скоростью четыре оборота в секунду, а само подобное вращение происходит дискретными скачками по 120°, каждый из которых сопровождается образованием одной молекулы АТФ.
Непосредственная функция синтеза АТФ локализована на β-субъединицах сопрягающего комплекса F 1 . При этом самым первым актом в цепи событий, приводящих к образованию АТФ, является связывание АДФ и фосфата с активным центром свободной β-субъединицы, находящейся в состоянии 1. За счёт энергии внешнего источника (тока протонов) в комплексе F 1 происходят конформационные изменения, в результате которых АДФ и фосфат становятся прочно связанными с каталитическим центром (состояние 2), где становится возможным образование ковалентной связи между ними, ведущей к образованию АТФ. На данной стадии АТФ-синтазы ферменту практически не требуется энергии, которая будет необходима на следующем этапе для освобождения прочно связанной молекулы АТФ из ферментативного центра. Поэтому следующий этап работы фермента заключается в том, чтобы в результате энергозависимого структурного изменения комплекса F 1 каталитическая β-субъединица, содержащая прочно связанную молекулу АТФ, перешла в состояние 3, в котором связь АТФ с каталитическим центром ослаблена. В результате этого молекула АТФ покидает фермент, а β-субъединица возвращается в исходное состояние 1, благодаря чему обеспечивается цикличность работы фермента.
Работа АТФ-синтазы связана с механическими движениями её отдельных частей, что позволило отнести этот процесс к особому типу явлений, названных «вращательным катализом». Подобно тому, как электрический ток в обмотке электродвигателя приводит в движение ротор относительно статора, направленный перенос протонов через АТФ-синтазу вызывает вращение отдельных субъединиц фактора сопряжения F 1 относительно других субъединиц ферментного комплекса, в результате чего это уникальное энергообразующее устройство совершает химическую работу - синтезирует молекулы АТФ. В дальнейшем АТФ поступает в цитоплазму клетки, где расходуется на самые разнообразные энергозависимые процессы. Подобный перенос осуществляется специальным встроенным в мембрану митохондрий ферментом АТФ/АДФ-транслоказой, который обменивает вновь синтезированную АТФ на цитоплазматическую АДФ, что гарантирует сохранность фонда адениловых нуклеотидов внутри митохондрий.
Митохондрии и наследственность
ДНК митохондрий наследуются почти исключительно по материнской линии. Каждая митохондрия имеет несколько участков нуклеотидов в ДНК, идентичных во всех митохондриях (то есть в клетке много копий митохондриальных ДНК), что очень важно для митохондрий, неспособных восстанавливать ДНК от повреждений (наблюдается высокая частота мутаций). Мутации в митохондриальной ДНК являются причиной целого ряда наследственных заболеваний человека.
См. также
Напишите отзыв о статье "Митохондрия"
Примечания
Литература
- М. Б. Беркинблит, С. М. Глаголев, В. А. Фуралев . Общая биология. - М.: МИРОС, 1999.
- Д. Тейлор, Н. Грин, У. Стаут . Биология. - М.: МИР, 2006.
- Э. Уиллет . Генетика без тайн. - М.: ЭКСМО, 2008.
- Д. Г. Дерябин . Функциональная морфология клетки. - М.: КДУ, 2005.
- Белякович А.Г. Изучение митохондрий и бактерий с помощью соли тетразолия п-НТФ. - Пущино: ОНТИ НЦБИ АН СССР, 1990.
- Н. Л. Векшин. Флуоресцентная спектроскопия биополимеров. Пущино, Фотон, 2009.
Ссылки
- Ченцов Ю. С. , 1997
|
||||||||||||||||||||||
Отрывок, характеризующий Митохондрия
Платону Каратаеву должно было быть за пятьдесят лет, судя по его рассказам о походах, в которых он участвовал давнишним солдатом. Он сам не знал и никак не мог определить, сколько ему было лет; но зубы его, ярко белые и крепкие, которые все выкатывались своими двумя полукругами, когда он смеялся (что он часто делал), были все хороши и целы; ни одного седого волоса не было в его бороде и волосах, и все тело его имело вид гибкости и в особенности твердости и сносливости.Лицо его, несмотря на мелкие круглые морщинки, имело выражение невинности и юности; голос у него был приятный и певучий. Но главная особенность его речи состояла в непосредственности и спорости. Он, видимо, никогда не думал о том, что он сказал и что он скажет; и от этого в быстроте и верности его интонаций была особенная неотразимая убедительность.
Физические силы его и поворотливость были таковы первое время плена, что, казалось, он не понимал, что такое усталость и болезнь. Каждый день утром а вечером он, ложась, говорил: «Положи, господи, камушком, подними калачиком»; поутру, вставая, всегда одинаково пожимая плечами, говорил: «Лег – свернулся, встал – встряхнулся». И действительно, стоило ему лечь, чтобы тотчас же заснуть камнем, и стоило встряхнуться, чтобы тотчас же, без секунды промедления, взяться за какое нибудь дело, как дети, вставши, берутся за игрушки. Он все умел делать, не очень хорошо, но и не дурно. Он пек, парил, шил, строгал, тачал сапоги. Он всегда был занят и только по ночам позволял себе разговоры, которые он любил, и песни. Он пел песни, не так, как поют песенники, знающие, что их слушают, но пел, как поют птицы, очевидно, потому, что звуки эти ему было так же необходимо издавать, как необходимо бывает потянуться или расходиться; и звуки эти всегда бывали тонкие, нежные, почти женские, заунывные, и лицо его при этом бывало очень серьезно.
Попав в плен и обросши бородою, он, видимо, отбросил от себя все напущенное на него, чуждое, солдатское и невольно возвратился к прежнему, крестьянскому, народному складу.
– Солдат в отпуску – рубаха из порток, – говаривал он. Он неохотно говорил про свое солдатское время, хотя не жаловался, и часто повторял, что он всю службу ни разу бит не был. Когда он рассказывал, то преимущественно рассказывал из своих старых и, видимо, дорогих ему воспоминаний «христианского», как он выговаривал, крестьянского быта. Поговорки, которые наполняли его речь, не были те, большей частью неприличные и бойкие поговорки, которые говорят солдаты, но это были те народные изречения, которые кажутся столь незначительными, взятые отдельно, и которые получают вдруг значение глубокой мудрости, когда они сказаны кстати.
Часто он говорил совершенно противоположное тому, что он говорил прежде, но и то и другое было справедливо. Он любил говорить и говорил хорошо, украшая свою речь ласкательными и пословицами, которые, Пьеру казалось, он сам выдумывал; но главная прелесть его рассказов состояла в том, что в его речи события самые простые, иногда те самые, которые, не замечая их, видел Пьер, получали характер торжественного благообразия. Он любил слушать сказки, которые рассказывал по вечерам (всё одни и те же) один солдат, но больше всего он любил слушать рассказы о настоящей жизни. Он радостно улыбался, слушая такие рассказы, вставляя слова и делая вопросы, клонившиеся к тому, чтобы уяснить себе благообразие того, что ему рассказывали. Привязанностей, дружбы, любви, как понимал их Пьер, Каратаев не имел никаких; но он любил и любовно жил со всем, с чем его сводила жизнь, и в особенности с человеком – не с известным каким нибудь человеком, а с теми людьми, которые были перед его глазами. Он любил свою шавку, любил товарищей, французов, любил Пьера, который был его соседом; но Пьер чувствовал, что Каратаев, несмотря на всю свою ласковую нежность к нему (которою он невольно отдавал должное духовной жизни Пьера), ни на минуту не огорчился бы разлукой с ним. И Пьер то же чувство начинал испытывать к Каратаеву.
Платон Каратаев был для всех остальных пленных самым обыкновенным солдатом; его звали соколик или Платоша, добродушно трунили над ним, посылали его за посылками. Но для Пьера, каким он представился в первую ночь, непостижимым, круглым и вечным олицетворением духа простоты и правды, таким он и остался навсегда.
Платон Каратаев ничего не знал наизусть, кроме своей молитвы. Когда он говорил свои речи, он, начиная их, казалось, не знал, чем он их кончит.
Когда Пьер, иногда пораженный смыслом его речи, просил повторить сказанное, Платон не мог вспомнить того, что он сказал минуту тому назад, – так же, как он никак не мог словами сказать Пьеру свою любимую песню. Там было: «родимая, березанька и тошненько мне», но на словах не выходило никакого смысла. Он не понимал и не мог понять значения слов, отдельно взятых из речи. Каждое слово его и каждое действие было проявлением неизвестной ему деятельности, которая была его жизнь. Но жизнь его, как он сам смотрел на нее, не имела смысла как отдельная жизнь. Она имела смысл только как частица целого, которое он постоянно чувствовал. Его слова и действия выливались из него так же равномерно, необходимо и непосредственно, как запах отделяется от цветка. Он не мог понять ни цены, ни значения отдельно взятого действия или слова.
Получив от Николая известие о том, что брат ее находится с Ростовыми, в Ярославле, княжна Марья, несмотря на отговариванья тетки, тотчас же собралась ехать, и не только одна, но с племянником. Трудно ли, нетрудно, возможно или невозможно это было, она не спрашивала и не хотела знать: ее обязанность была не только самой быть подле, может быть, умирающего брата, но и сделать все возможное для того, чтобы привезти ему сына, и она поднялась ехать. Если князь Андрей сам не уведомлял ее, то княжна Марья объясняла ото или тем, что он был слишком слаб, чтобы писать, или тем, что он считал для нее и для своего сына этот длинный переезд слишком трудным и опасным.
В несколько дней княжна Марья собралась в дорогу. Экипажи ее состояли из огромной княжеской кареты, в которой она приехала в Воронеж, брички и повозки. С ней ехали m lle Bourienne, Николушка с гувернером, старая няня, три девушки, Тихон, молодой лакей и гайдук, которого тетка отпустила с нею.
Ехать обыкновенным путем на Москву нельзя было и думать, и потому окольный путь, который должна была сделать княжна Марья: на Липецк, Рязань, Владимир, Шую, был очень длинен, по неимению везде почтовых лошадей, очень труден и около Рязани, где, как говорили, показывались французы, даже опасен.
Во время этого трудного путешествия m lle Bourienne, Десаль и прислуга княжны Марьи были удивлены ее твердостью духа и деятельностью. Она позже всех ложилась, раньше всех вставала, и никакие затруднения не могли остановить ее. Благодаря ее деятельности и энергии, возбуждавшим ее спутников, к концу второй недели они подъезжали к Ярославлю.
В последнее время своего пребывания в Воронеже княжна Марья испытала лучшее счастье в своей жизни. Любовь ее к Ростову уже не мучила, не волновала ее. Любовь эта наполняла всю ее душу, сделалась нераздельною частью ее самой, и она не боролась более против нее. В последнее время княжна Марья убедилась, – хотя она никогда ясно словами определенно не говорила себе этого, – убедилась, что она была любима и любила. В этом она убедилась в последнее свое свидание с Николаем, когда он приехал ей объявить о том, что ее брат был с Ростовыми. Николай ни одним словом не намекнул на то, что теперь (в случае выздоровления князя Андрея) прежние отношения между ним и Наташей могли возобновиться, но княжна Марья видела по его лицу, что он знал и думал это. И, несмотря на то, его отношения к ней – осторожные, нежные и любовные – не только не изменились, но он, казалось, радовался тому, что теперь родство между ним и княжной Марьей позволяло ему свободнее выражать ей свою дружбу любовь, как иногда думала княжна Марья. Княжна Марья знала, что она любила в первый и последний раз в жизни, и чувствовала, что она любима, и была счастлива, спокойна в этом отношении.
Но это счастье одной стороны душевной не только не мешало ей во всей силе чувствовать горе о брате, но, напротив, это душевное спокойствие в одном отношении давало ей большую возможность отдаваться вполне своему чувству к брату. Чувство это было так сильно в первую минуту выезда из Воронежа, что провожавшие ее были уверены, глядя на ее измученное, отчаянное лицо, что она непременно заболеет дорогой; но именно трудности и заботы путешествия, за которые с такою деятельностью взялась княжна Марья, спасли ее на время от ее горя и придали ей силы.
Как и всегда это бывает во время путешествия, княжна Марья думала только об одном путешествии, забывая о том, что было его целью. Но, подъезжая к Ярославлю, когда открылось опять то, что могло предстоять ей, и уже не через много дней, а нынче вечером, волнение княжны Марьи дошло до крайних пределов.
Когда посланный вперед гайдук, чтобы узнать в Ярославле, где стоят Ростовы и в каком положении находится князь Андрей, встретил у заставы большую въезжавшую карету, он ужаснулся, увидав страшно бледное лицо княжны, которое высунулось ему из окна.
– Все узнал, ваше сиятельство: ростовские стоят на площади, в доме купца Бронникова. Недалече, над самой над Волгой, – сказал гайдук.
Княжна Марья испуганно вопросительно смотрела на его лицо, не понимая того, что он говорил ей, не понимая, почему он не отвечал на главный вопрос: что брат? M lle Bourienne сделала этот вопрос за княжну Марью.
– Что князь? – спросила она.
– Их сиятельство с ними в том же доме стоят.
«Стало быть, он жив», – подумала княжна и тихо спросила: что он?
– Люди сказывали, все в том же положении.
Что значило «все в том же положении», княжна не стала спрашивать и мельком только, незаметно взглянув на семилетнего Николушку, сидевшего перед нею и радовавшегося на город, опустила голову и не поднимала ее до тех пор, пока тяжелая карета, гремя, трясясь и колыхаясь, не остановилась где то. Загремели откидываемые подножки.
Отворились дверцы. Слева была вода – река большая, справа было крыльцо; на крыльце были люди, прислуга и какая то румяная, с большой черной косой, девушка, которая неприятно притворно улыбалась, как показалось княжне Марье (это была Соня). Княжна взбежала по лестнице, притворно улыбавшаяся девушка сказала: – Сюда, сюда! – и княжна очутилась в передней перед старой женщиной с восточным типом лица, которая с растроганным выражением быстро шла ей навстречу. Это была графиня. Она обняла княжну Марью и стала целовать ее.
– Mon enfant! – проговорила она, – je vous aime et vous connais depuis longtemps. [Дитя мое! я вас люблю и знаю давно.]
Несмотря на все свое волнение, княжна Марья поняла, что это была графиня и что надо было ей сказать что нибудь. Она, сама не зная как, проговорила какие то учтивые французские слова, в том же тоне, в котором были те, которые ей говорили, и спросила: что он?
– Доктор говорит, что нет опасности, – сказала графиня, но в то время, как она говорила это, она со вздохом подняла глаза кверху, и в этом жесте было выражение, противоречащее ее словам.
– Где он? Можно его видеть, можно? – спросила княжна.
– Сейчас, княжна, сейчас, мой дружок. Это его сын? – сказала она, обращаясь к Николушке, который входил с Десалем. – Мы все поместимся, дом большой. О, какой прелестный мальчик!
Графиня ввела княжну в гостиную. Соня разговаривала с m lle Bourienne. Графиня ласкала мальчика. Старый граф вошел в комнату, приветствуя княжну. Старый граф чрезвычайно переменился с тех пор, как его последний раз видела княжна. Тогда он был бойкий, веселый, самоуверенный старичок, теперь он казался жалким, затерянным человеком. Он, говоря с княжной, беспрестанно оглядывался, как бы спрашивая у всех, то ли он делает, что надобно. После разорения Москвы и его имения, выбитый из привычной колеи, он, видимо, потерял сознание своего значения и чувствовал, что ему уже нет места в жизни.
Несмотря на то волнение, в котором она находилась, несмотря на одно желание поскорее увидать брата и на досаду за то, что в эту минуту, когда ей одного хочется – увидать его, – ее занимают и притворно хвалят ее племянника, княжна замечала все, что делалось вокруг нее, и чувствовала необходимость на время подчиниться этому новому порядку, в который она вступала. Она знала, что все это необходимо, и ей было это трудно, но она не досадовала на них.
– Это моя племянница, – сказал граф, представляя Соню, – вы не знаете ее, княжна?
Княжна повернулась к ней и, стараясь затушить поднявшееся в ее душе враждебное чувство к этой девушке, поцеловала ее. Но ей становилось тяжело оттого, что настроение всех окружающих было так далеко от того, что было в ее душе.
– Где он? – спросила она еще раз, обращаясь ко всем.
– Он внизу, Наташа с ним, – отвечала Соня, краснея. – Пошли узнать. Вы, я думаю, устали, княжна?
У княжны выступили на глаза слезы досады. Она отвернулась и хотела опять спросить у графини, где пройти к нему, как в дверях послышались легкие, стремительные, как будто веселые шаги. Княжна оглянулась и увидела почти вбегающую Наташу, ту Наташу, которая в то давнишнее свидание в Москве так не понравилась ей.
Но не успела княжна взглянуть на лицо этой Наташи, как она поняла, что это был ее искренний товарищ по горю, и потому ее друг. Она бросилась ей навстречу и, обняв ее, заплакала на ее плече.
Как только Наташа, сидевшая у изголовья князя Андрея, узнала о приезде княжны Марьи, она тихо вышла из его комнаты теми быстрыми, как показалось княжне Марье, как будто веселыми шагами и побежала к ней.
На взволнованном лице ее, когда она вбежала в комнату, было только одно выражение – выражение любви, беспредельной любви к нему, к ней, ко всему тому, что было близко любимому человеку, выраженье жалости, страданья за других и страстного желанья отдать себя всю для того, чтобы помочь им. Видно было, что в эту минуту ни одной мысли о себе, о своих отношениях к нему не было в душе Наташи.
Чуткая княжна Марья с первого взгляда на лицо Наташи поняла все это и с горестным наслаждением плакала на ее плече.
– Пойдемте, пойдемте к нему, Мари, – проговорила Наташа, отводя ее в другую комнату.
Княжна Марья подняла лицо, отерла глаза и обратилась к Наташе. Она чувствовала, что от нее она все поймет и узнает.
– Что… – начала она вопрос, но вдруг остановилась. Она почувствовала, что словами нельзя ни спросить, ни ответить. Лицо и глаза Наташи должны были сказать все яснее и глубже.
Наташа смотрела на нее, но, казалось, была в страхе и сомнении – сказать или не сказать все то, что она знала; она как будто почувствовала, что перед этими лучистыми глазами, проникавшими в самую глубь ее сердца, нельзя не сказать всю, всю истину, какою она ее видела. Губа Наташи вдруг дрогнула, уродливые морщины образовались вокруг ее рта, и она, зарыдав, закрыла лицо руками.
Княжна Марья поняла все.
Но она все таки надеялась и спросила словами, в которые она не верила:
– Но как его рана? Вообще в каком он положении?
– Вы, вы… увидите, – только могла сказать Наташа.
Они посидели несколько времени внизу подле его комнаты, с тем чтобы перестать плакать и войти к нему с спокойными лицами.
– Как шла вся болезнь? Давно ли ему стало хуже? Когда это случилось? – спрашивала княжна Марья.
Наташа рассказывала, что первое время была опасность от горячечного состояния и от страданий, но в Троице это прошло, и доктор боялся одного – антонова огня. Но и эта опасность миновалась. Когда приехали в Ярославль, рана стала гноиться (Наташа знала все, что касалось нагноения и т. п.), и доктор говорил, что нагноение может пойти правильно. Сделалась лихорадка. Доктор говорил, что лихорадка эта не так опасна.
– Но два дня тому назад, – начала Наташа, – вдруг это сделалось… – Она удержала рыданья. – Я не знаю отчего, но вы увидите, какой он стал.
– Ослабел? похудел?.. – спрашивала княжна.
– Нет, не то, но хуже. Вы увидите. Ах, Мари, Мари, он слишком хорош, он не может, не может жить… потому что…
Когда Наташа привычным движением отворила его дверь, пропуская вперед себя княжну, княжна Марья чувствовала уже в горле своем готовые рыданья. Сколько она ни готовилась, ни старалась успокоиться, она знала, что не в силах будет без слез увидать его.
Княжна Марья понимала то, что разумела Наташа словами: сним случилось это два дня тому назад. Она понимала, что это означало то, что он вдруг смягчился, и что смягчение, умиление эти были признаками смерти. Она, подходя к двери, уже видела в воображении своем то лицо Андрюши, которое она знала с детства, нежное, кроткое, умиленное, которое так редко бывало у него и потому так сильно всегда на нее действовало. Она знала, что он скажет ей тихие, нежные слова, как те, которые сказал ей отец перед смертью, и что она не вынесет этого и разрыдается над ним. Но, рано ли, поздно ли, это должно было быть, и она вошла в комнату. Рыдания все ближе и ближе подступали ей к горлу, в то время как она своими близорукими глазами яснее и яснее различала его форму и отыскивала его черты, и вот она увидала его лицо и встретилась с ним взглядом.
Он лежал на диване, обложенный подушками, в меховом беличьем халате. Он был худ и бледен. Одна худая, прозрачно белая рука его держала платок, другою он, тихими движениями пальцев, трогал тонкие отросшие усы. Глаза его смотрели на входивших.
Увидав его лицо и встретившись с ним взглядом, княжна Марья вдруг умерила быстроту своего шага и почувствовала, что слезы вдруг пересохли и рыдания остановились. Уловив выражение его лица и взгляда, она вдруг оробела и почувствовала себя виноватой.
«Да в чем же я виновата?» – спросила она себя. «В том, что живешь и думаешь о живом, а я!..» – отвечал его холодный, строгий взгляд.
В глубоком, не из себя, но в себя смотревшем взгляде была почти враждебность, когда он медленно оглянул сестру и Наташу.
Он поцеловался с сестрой рука в руку, по их привычке.
– Здравствуй, Мари, как это ты добралась? – сказал он голосом таким же ровным и чуждым, каким был его взгляд. Ежели бы он завизжал отчаянным криком, то этот крик менее бы ужаснул княжну Марью, чем звук этого голоса.
– И Николушку привезла? – сказал он также ровно и медленно и с очевидным усилием воспоминанья.
– Как твое здоровье теперь? – говорила княжна Марья, сама удивляясь тому, что она говорила.
– Это, мой друг, у доктора спрашивать надо, – сказал он, и, видимо сделав еще усилие, чтобы быть ласковым, он сказал одним ртом (видно было, что он вовсе не думал того, что говорил): – Merci, chere amie, d"etre venue. [Спасибо, милый друг, что приехала.]
Княжна Марья пожала его руку. Он чуть заметно поморщился от пожатия ее руки. Он молчал, и она не знала, что говорить. Она поняла то, что случилось с ним за два дня. В словах, в тоне его, в особенности во взгляде этом – холодном, почти враждебном взгляде – чувствовалась страшная для живого человека отчужденность от всего мирского. Он, видимо, с трудом понимал теперь все живое; но вместе с тем чувствовалось, что он не понимал живого не потому, чтобы он был лишен силы понимания, но потому, что он понимал что то другое, такое, чего не понимали и не могли понять живые и что поглощало его всего.
– Да, вот как странно судьба свела нас! – сказал он, прерывая молчание и указывая на Наташу. – Она все ходит за мной.
Княжна Марья слушала и не понимала того, что он говорил. Он, чуткий, нежный князь Андрей, как мог он говорить это при той, которую он любил и которая его любила! Ежели бы он думал жить, то не таким холодно оскорбительным тоном он сказал бы это. Ежели бы он не знал, что умрет, то как же ему не жалко было ее, как он мог при ней говорить это! Одно объяснение только могло быть этому, это то, что ему было все равно, и все равно оттого, что что то другое, важнейшее, было открыто ему.
Разговор был холодный, несвязный и прерывался беспрестанно.
– Мари проехала через Рязань, – сказала Наташа. Князь Андрей не заметил, что она называла его сестру Мари. А Наташа, при нем назвав ее так, в первый раз сама это заметила.
– Ну что же? – сказал он.
– Ей рассказывали, что Москва вся сгорела, совершенно, что будто бы…
Наташа остановилась: нельзя было говорить. Он, очевидно, делал усилия, чтобы слушать, и все таки не мог.
– Да, сгорела, говорят, – сказал он. – Это очень жалко, – и он стал смотреть вперед, пальцами рассеянно расправляя усы.
– А ты встретилась с графом Николаем, Мари? – сказал вдруг князь Андрей, видимо желая сделать им приятное. – Он писал сюда, что ты ему очень полюбилась, – продолжал он просто, спокойно, видимо не в силах понимать всего того сложного значения, которое имели его слова для живых людей. – Ежели бы ты его полюбила тоже, то было бы очень хорошо… чтобы вы женились, – прибавил он несколько скорее, как бы обрадованный словами, которые он долго искал и нашел наконец. Княжна Марья слышала его слова, но они не имели для нее никакого другого значения, кроме того, что они доказывали то, как страшно далек он был теперь от всего живого.
– Что обо мне говорить! – сказала она спокойно и взглянула на Наташу. Наташа, чувствуя на себе ее взгляд, не смотрела на нее. Опять все молчали.
– Andre, ты хоч… – вдруг сказала княжна Марья содрогнувшимся голосом, – ты хочешь видеть Николушку? Он все время вспоминал о тебе.
Князь Андрей чуть заметно улыбнулся в первый раз, но княжна Марья, так знавшая его лицо, с ужасом поняла, что это была улыбка не радости, не нежности к сыну, но тихой, кроткой насмешки над тем, что княжна Марья употребляла, по ее мнению, последнее средство для приведения его в чувства.
– Да, я очень рад Николушке. Он здоров?
Когда привели к князю Андрею Николушку, испуганно смотревшего на отца, но не плакавшего, потому что никто не плакал, князь Андрей поцеловал его и, очевидно, не знал, что говорить с ним.
Когда Николушку уводили, княжна Марья подошла еще раз к брату, поцеловала его и, не в силах удерживаться более, заплакала.
Он пристально посмотрел на нее.
– Ты об Николушке? – сказал он.
Княжна Марья, плача, утвердительно нагнула голову.
– Мари, ты знаешь Еван… – но он вдруг замолчал.
– Что ты говоришь?
– Ничего. Не надо плакать здесь, – сказал он, тем же холодным взглядом глядя на нее.
Когда княжна Марья заплакала, он понял, что она плакала о том, что Николушка останется без отца. С большим усилием над собой он постарался вернуться назад в жизнь и перенесся на их точку зрения.
«Да, им это должно казаться жалко! – подумал он. – А как это просто!»
«Птицы небесные ни сеют, ни жнут, но отец ваш питает их», – сказал он сам себе и хотел то же сказать княжне. «Но нет, они поймут это по своему, они не поймут! Этого они не могут понимать, что все эти чувства, которыми они дорожат, все наши, все эти мысли, которые кажутся нам так важны, что они – не нужны. Мы не можем понимать друг друга». – И он замолчал.
Маленькому сыну князя Андрея было семь лет. Он едва умел читать, он ничего не знал. Он многое пережил после этого дня, приобретая знания, наблюдательность, опытность; но ежели бы он владел тогда всеми этими после приобретенными способностями, он не мог бы лучше, глубже понять все значение той сцены, которую он видел между отцом, княжной Марьей и Наташей, чем он ее понял теперь. Он все понял и, не плача, вышел из комнаты, молча подошел к Наташе, вышедшей за ним, застенчиво взглянул на нее задумчивыми прекрасными глазами; приподнятая румяная верхняя губа его дрогнула, он прислонился к ней головой и заплакал.
С этого дня он избегал Десаля, избегал ласкавшую его графиню и либо сидел один, либо робко подходил к княжне Марье и к Наташе, которую он, казалось, полюбил еще больше своей тетки, и тихо и застенчиво ласкался к ним.
Княжна Марья, выйдя от князя Андрея, поняла вполне все то, что сказало ей лицо Наташи. Она не говорила больше с Наташей о надежде на спасение его жизни. Она чередовалась с нею у его дивана и не плакала больше, но беспрестанно молилась, обращаясь душою к тому вечному, непостижимому, которого присутствие так ощутительно было теперь над умиравшим человеком.
Князь Андрей не только знал, что он умрет, но он чувствовал, что он умирает, что он уже умер наполовину. Он испытывал сознание отчужденности от всего земного и радостной и странной легкости бытия. Он, не торопясь и не тревожась, ожидал того, что предстояло ему. То грозное, вечное, неведомое и далекое, присутствие которого он не переставал ощущать в продолжение всей своей жизни, теперь для него было близкое и – по той странной легкости бытия, которую он испытывал, – почти понятное и ощущаемое.
Прежде он боялся конца. Он два раза испытал это страшное мучительное чувство страха смерти, конца, и теперь уже не понимал его.
Первый раз он испытал это чувство тогда, когда граната волчком вертелась перед ним и он смотрел на жнивье, на кусты, на небо и знал, что перед ним была смерть. Когда он очнулся после раны и в душе его, мгновенно, как бы освобожденный от удерживавшего его гнета жизни, распустился этот цветок любви, вечной, свободной, не зависящей от этой жизни, он уже не боялся смерти и не думал о ней.
Чем больше он, в те часы страдальческого уединения и полубреда, которые он провел после своей раны, вдумывался в новое, открытое ему начало вечной любви, тем более он, сам не чувствуя того, отрекался от земной жизни. Всё, всех любить, всегда жертвовать собой для любви, значило никого не любить, значило не жить этою земною жизнию. И чем больше он проникался этим началом любви, тем больше он отрекался от жизни и тем совершеннее уничтожал ту страшную преграду, которая без любви стоит между жизнью и смертью. Когда он, это первое время, вспоминал о том, что ему надо было умереть, он говорил себе: ну что ж, тем лучше.
Но после той ночи в Мытищах, когда в полубреду перед ним явилась та, которую он желал, и когда он, прижав к своим губам ее руку, заплакал тихими, радостными слезами, любовь к одной женщине незаметно закралась в его сердце и опять привязала его к жизни. И радостные и тревожные мысли стали приходить ему. Вспоминая ту минуту на перевязочном пункте, когда он увидал Курагина, он теперь не мог возвратиться к тому чувству: его мучил вопрос о том, жив ли он? И он не смел спросить этого.
Болезнь его шла своим физическим порядком, но то, что Наташа называла: это сделалось с ним, случилось с ним два дня перед приездом княжны Марьи. Это была та последняя нравственная борьба между жизнью и смертью, в которой смерть одержала победу. Это было неожиданное сознание того, что он еще дорожил жизнью, представлявшейся ему в любви к Наташе, и последний, покоренный припадок ужаса перед неведомым.
Это было вечером. Он был, как обыкновенно после обеда, в легком лихорадочном состоянии, и мысли его были чрезвычайно ясны. Соня сидела у стола. Он задремал. Вдруг ощущение счастья охватило его.
«А, это она вошла!» – подумал он.
Действительно, на месте Сони сидела только что неслышными шагами вошедшая Наташа.
С тех пор как она стала ходить за ним, он всегда испытывал это физическое ощущение ее близости. Она сидела на кресле, боком к нему, заслоняя собой от него свет свечи, и вязала чулок. (Она выучилась вязать чулки с тех пор, как раз князь Андрей сказал ей, что никто так не умеет ходить за больными, как старые няни, которые вяжут чулки, и что в вязании чулка есть что то успокоительное.) Тонкие пальцы ее быстро перебирали изредка сталкивающиеся спицы, и задумчивый профиль ее опущенного лица был ясно виден ему. Она сделала движенье – клубок скатился с ее колен. Она вздрогнула, оглянулась на него и, заслоняя свечу рукой, осторожным, гибким и точным движением изогнулась, подняла клубок и села в прежнее положение.
Он смотрел на нее, не шевелясь, и видел, что ей нужно было после своего движения вздохнуть во всю грудь, но она не решалась этого сделать и осторожно переводила дыханье.
В Троицкой лавре они говорили о прошедшем, и он сказал ей, что, ежели бы он был жив, он бы благодарил вечно бога за свою рану, которая свела его опять с нею; но с тех пор они никогда не говорили о будущем.
«Могло или не могло это быть? – думал он теперь, глядя на нее и прислушиваясь к легкому стальному звуку спиц. – Неужели только затем так странно свела меня с нею судьба, чтобы мне умереть?.. Неужели мне открылась истина жизни только для того, чтобы я жил во лжи? Я люблю ее больше всего в мире. Но что же делать мне, ежели я люблю ее?» – сказал он, и он вдруг невольно застонал, по привычке, которую он приобрел во время своих страданий.
Услыхав этот звук, Наташа положила чулок, перегнулась ближе к нему и вдруг, заметив его светящиеся глаза, подошла к нему легким шагом и нагнулась.
– Вы не спите?
– Нет, я давно смотрю на вас; я почувствовал, когда вы вошли. Никто, как вы, но дает мне той мягкой тишины… того света. Мне так и хочется плакать от радости.
II. Митохондрии (строение и функции)
Полисомы. Синтез цитоплазматических белков
Рибосомы представляют собой мельчайшие органеллы, присутствующие в цитоплазме клетки. Несмотря на свои размеры, они являются сложными молекулярными ансамблеями, состоящими из рибосомальной РНК (р-РНК) различной длины и рибосомальных белков . В цитоплазме рибосомы встречаются в виде 2-х форм:
1. В диссоциированном состоянии (две субъединицы: малая и большая), которое свидетельствует об их неактивном статусе;
2. В ассоциированном виде – это форма их активного статуса.
Большая субъединица образуется тремя молекулами РНК, имеет форму полушара с 3 выступами, взаимодействующие с «шипиками» малой субъединицы.
Малая субъединица содержит лишь одну молекулу РНК и выглядит в виде «шапочки» с шипиками, обращёнными в сторону большой субъединицы. Ассоциация субъединиц рибосомы – это взаимодействие рельефов их поверхностей.
Функции субъединиц:
1. Малая ответственна за связывание с матричной РНК;
2. Большая – за образование полипептидной цепи.
Полисомы – это группа рибосом (от 5 до 30) связанных нитью м-РНК с образованием функционального комплекса. На нём происходит синтез цитоплазматических белков, необходимых клетке для роста, развития органелл дифференцировки.
Этапы синтеза цитоплазматических белков:
1. Выход из ядра м-РНК;
2. Сборка рибосом;
3. Образование функциональной полисомы;
4. Синтез сигнального пептида;
5. Считывание последовательности аминокислот в составе пептида сигнал-распознающей частицы (СРЧ);
6. Завершение синтеза цитоплазматического белка на полисоме. См рис. 1
Рис. 1: Схема синтеза цитоплазматических белков
II. Митохондрии (строение и функции)
Митохондрии – это система энергообеспечения клетки. На светооптическом уровне их выявляют при окраске по Альтману, они выглядят в виде зёрнышек и нитей. В цитоплазме они распределены диффузно, а в специализированных клетках сосредоточенны в участках, где имеется наибольшая потребность в энергии.
 Электронномикроскопический уровень организации митохондрии
: в ней выделяют две мембраны: наружную и внутреннюю. См. рис. 2
Электронномикроскопический уровень организации митохондрии
: в ней выделяют две мембраны: наружную и внутреннюю. См. рис. 2
Рис. 2: Схема строения митохондрии
Наружная мембрана – это мешок с относительно ровной поверхностью, она по химическому составу и свойствам близка к плазмолемме, отличается она более высокой проницаемостью и содержит ферменты метаболизма жирных кислот, фосфолипидов и липидов.
Функция:
1. Отграничение митохондрии в гиалоплазме;
2. Транспорт в митохондрию субстратов для клеточного дыхания.
Внутренняя мембрана – неровная, она формирует кристы в виде пластин (ламеллярные кристы) с увеличением площади её поверхности. Главным компонентом этой мембраны являются молекулы белков, относящиеся к ферментам дыхательной цепи, цитохромы.
На поверхности крист в некоторых клетках описывают грибовидные частицы (F 1 -частицы), в которых различают головку (9 нм) и ножку (3 нм). Считают, что именно здесь происходит синтез АТФ и АДФ.
Между наружной и внутренней мембранами образуется небольшое (около 15 – 20 нм) пространство, которое называют наружной камерой митохондрий. Внутренняя камера ограничена соответственно внутренней митохондриальной мембраной и содержит матрикс.
Матрикс митохондрий имеет гелеобразную фазу и отличается высоким содержанием белка. В нём встречаются митохондриальные гранулы – частицы диаметром 20 – 50 нм высокой электронной плотности, они содержат ионы Са 2+ и Mg 2+ . Матрикс митохондрий содержит также митохондриальные ДНК и рибосомы. На первых происходит синтез транспортных белков митохондриальных мембран и некоторых белков, участвующих в фосфолировании АДФ. ДНК здесь состоит из 37 генов и не содержит некодирующие последовательность нуклеотидов.
Функции митохондрий:
1. Обеспечение клетки энергией в виде АТФ;
2. Участие в синтезе стероидных гормонов;
3. Участие в синтезе нуклеиновых кислот;
4. Депонирование кальция.
Митохондрии – строение и функции
| Наименование параметра | Значение |
| Тема статьи: | Митохондрии – строение и функции |
| Рубрика (тематическая категория) | Экология |
Общая морфология. Митохондрии или хондриосомы (от греч. mitos– нить, chondrion- зернышко, soma- тельце) представляют из себягранулярные или нитевидные органеллы, присутствующие в цитоплазме простейших, растений и животных (рис. 198). Митохондрии можно наблюдать в живых клетках, так как они обладают достаточно высокой плотностью. В живых клетках митохондрии могут двигаться, перемещаться, сливаться друг с другом.
У разных видов размеры митохондрий очень непостоянны, так же как изменчива их форма (рис. 199). Все же у большинства клеток толщина этих структур относительно постоянна (около 0,5 мкм), а длина колеблется, достигая у нитчатых форм до 7-60 мкм.
Изучение величины и числа митохондрий не такое простое дело. Это связано с тем, что размеры и число митохондрий, которые видны на ультратонких срезах, не соответствуют реальности.
Обычные же подсчеты показывают, что на печеночную клетку приходится около 200 митохондрий. Это составляет более 20% от общего объёма цитоплазмы и около 30-35% от общего количества белка в клетке. Площадь поверхности всех митохондрий печеночной клетки в 4-5 раз больше поверхности ее плазматической мембраны. Больше всего митохондрий в ооцитах (около 300000) и у гигантской амебы Chaos chaos (до 500000).
В клетках зеленых растений число митохондрий меньше, чем в клетках животных, так как часть их функций могут выполнять хлоропласты.
Локализация митохондрии в клетках различна. Обычно митохондрии скапливаются вблизи тех участков цитоплазмы, где возникает потребность в АТФ, образующейся в митохондриях. Так, в скелетных мышцах митохондрии находятся вблизи миофибрилл. В сперматозоидах митохондрии образуют спиральный футляр вокруг оси жгутика; вероятно, это связано с крайне важно стью использования АТФ для движения хвоста сперматозоида. Аналогичным образом у простейших и в других клетках, снабженных ресничками, митохондрии локализуются непосредственно под клеточной мембраной у основания ресничек, для работы которых необходим АТФ. В аксонах нервных клеток митохондрии располагаются около синапсов, где происходит процесс передачи нервного импульса. В секреторных клетках, которые синтезируют большие количества белков, митохондрии тесно связаны с зонами эргастоплазмы; вероятно, они поставляют АТФ для активации аминокислот и синтеза белка на рибосомах.
Ультраструктура митохондрий. Митохондрии независимо от их величины или формы имеют универсальное строение, их ультраструктура однообразна. Митохондрии ограничены двумя мембранами (рис. 205). Наружняя митохондриальная мембрана отделяет ее от гиалоплазмы, она имеет ровные контуры, не образует впячиваний или складок, толщина составляет около 7 нм. На нее приходится около 7% от площади всех клеточных мембран. Мембрана не связана ни с какими другими мембранами цитоплазмы, замкнута сама на себя и представляет собой мембранный мешок. Наружнюю мембрану от внутренней отделяет межмембранное пространство шириной около 10-20 нм. Внутренняя мембрана (толщиной около 7 нм) ограничивает собственно внутреннее содержимое митохондрии, ее матрикс или митоплазму. Внутренняя мембрана митохондрий образовывает многочисленные впячивания внутрь митохондрий. Такие впячивания чаще всего имеют вид плоских гребней, или крист (рис. 206, 207а).
Общая поверхность внутренней мембраны митохондрии в печеночной клетке составляет примерно треть поверхности всех клеточных мембран. Митохондрии клеток сердечной мышцы содержат втрое больше крист, чем печеночные митохондрии что отражает различия в функциональных нагрузках митохондрий разных клеток. Расстояние между мембранами в кристе составляет около 10-20 нм.
Митохондриальные кристы, отходящие от внутренней мембраны и простирающиеся в сторону матрикса, не перегораживают полностью полость митохондрии и не нарушают непрерывности заполняющего ее матрикса.
Ориентация крист по отношению к длинной оси митохондрии различна для разных клеток. Ориентация должна быть перпендикулярная (клетки печени, почек) крист; в сердечной мышце наблюдается продольное расположение крист. Кристы могут ветвиться или образовывать пальцевидные отростки, изгибаться и не иметь выраженной ориентации (рис. 208). У простейших, одноклеточных водорослей, в некоторых клетках высших растений и животных выросты внутренней мембраны имеют вид трубок (трубчатые кристы).
Матрикс митохондрий имеет тонкозернистое гомогенное строение, в котором выявляются молекулы ДНК в виде тонких собранных в клубок нитей (около 2-3 нм) и митохондриальные рибосомы имеющие форму гранул размером около 15-20нм. Места отложения солей магния и кальция в матриксе образуют крупные (20-40 нм) плотные гранулы.
Функции митохондрий. Митохондрии осуществляют синтез АТФ, происходящий в результате процессов окисления органических субстратов и фосфорилирования АДФ.
Начальные этапы окисления углеводов называются анаэробным окислением, или гликолизом и происходят в гиалоплазме и не требуют участия кислорода. Субстратом окисления при анаэробном получении энергии служат гексозы и в первую очередь глюкоза; некоторые бактерии обладают свойством извлекать энергию, окисляяя пентозы, жирные кислоты или аминокислоты.
В глюкозе количество потенциальной энергии, заключенной в связях между атомами С, Н и О, составляет около 680 ккал на 1 моль (ᴛ.ᴇ. на 180 г глюкозы).
В живой клетке это огромное количество энергии освобождается в виде ступенчатого процесса, управляемого целым рядом окислительных ферментов, и не связано с переходом энергии химической связи в тепло, как при горении, а переходит в макроэнергетическую связь в молекулах АТФ, которые синтезируются при использовании освобождающейся энергии из АДФ и фосфата.
Образовавшиеся в результате гликолиза триозы, и в первую очередь пировиноградная кислота͵ в митохондриях вовлекаются в дальнейшее окисление. При этом происходит использование энергии расщепления всех химических связей, что приводит к выделению СО 2, к потреблению кислорода и синтезу большого количества АТФ. Эти процессы связаны с окислительным циклом трикарбоновых кислот и с дыхательной цепью переноса электронов, где происходит фосфорилирование АДФ и синтез клеточного “топлива”, молекул АТФ (рис. 209).
В цикле трикарбоновых кислот (цикл Кребса, или цикл лимонной кислоты) образовавшийся в результате гликолиза пируват сначала теряет молекулу СО 2 и, окисляясь до ацетата (двууглеродное соединение), соединяется с коферментом А. Далее ацетилкоэнзим А, соединяясь с оксалацетатом (четырехуглеродное соединение), образует шестиуглеродный цитрат (лимоную кислоту). Далее происходит цикл окисления этого шестиуглеродного соединения до четырехуглеродного оксалацетата͵ снова связывание с ацетилкоэнзимом А, и затем цикл повторяется. При этом окислении выделяются две молекулы СО 2, а электроны, освободившиеся при окислении, переносятся на акцепторные молекулы коферментов (NAD-никотинамидадениндинуклеотид), которые вовлекают их далее в цепь переноса электронов. Следовательно, в цикле трикарбоновых кислот нет самого синтеза АТФ, а идет окисление молекул, перенос электронов на акцепторы и выделение СО 2 . Все описанные выше события внутри митохондрий происходят в их матриксе.
Окисление исходного субстрата приводит к выделению СО 2 и воды, но при этом не выделяется тепловая энергия, как при горении, а образовываются молекулы АТФ. Οʜᴎ синтезируются другой группой белков, не связанных прямо с окислением. Во внутренних митохондриальных мембранах на поверхности мембран, смотрящих в матрикс, располагаются крупные белковые комплексы, ферменты, АТФ-синтетазы. В электронном микроскопе они видны в виде так называемых “грибовидных” телец сплошь выстилающие поверхность мембран, смотрящую в матрикс. Тельца имеют как бы ножку и головку, диаметром 8-9 нм. Следовательно, во внутренних мембранах митохондрий локализованы ферменты как окислительной цепи, так и ферменты синтеза АТФ (рис. 201б).
Дыхательная цепь - ϶ᴛᴏ главная система превращения энергии в митохондриях. Здесь происходит последовательное окисление и восстановление элементов дыхательной цепи, благодаря чему высвобождается небольшими порциями энергия. За счёт этой энергии в трех точках цепи из АДФ и фосфата образуется АТФ. По этой причине говорят, что окисление (перенос электронов) сопряжено с фосфорилированием (АДФ + Фн →АТФ, ᴛ.ᴇ. происходит процесс окислительного фосфорилирования.
Выделяющаяся при транспорте электронов энергия запасается в виде градиента протонов на мембране. Оказалось, что при переносе электронов в митохондриальной мембране каждый комплекс дыхательной цепи направляет свободную энергию окисление на перемещение протонов (положительных зарядов) через мембрану, из матрикса в межмембранное пространство, что приводит к образованию разности потенциалов на мембране: положительные заряды преобладают в межмембранном пространстве, а отрицательные – со стороны матрикса митохондрий. При достижении разности потенциалов (220 мВ) белковый комплекс АТФ-синтетазы начинает транспортировать протоны обратно в матрикс, при этом превращает одну форму энергии в другую: образует АТФ из АДФ и неорганического фосфата. Так происходит сопряжение окислительных процессов с синтетическим, с фосфорилированием АДФ. Пока происходит окисление субстратов, пока происходит перекачка протонов через внутреннюю митохондриальную мембрану – идет сопряженный с этим синтез АТФ, ᴛ.ᴇ. происходит окислительное фосфорилирование.
Эти два процесса можно разобщить. При этом продолжается перенос электронов, как и окисление субстрата͵ но синтеза АТФ не происходит. В этом случае энергия, освобождающаяся при окислении переходит в тепловую энергию.
Окислительное фосфорилирование у бактерий . У прокариотических клеток, способных к окислительному фосфорилированию, элементы цикла трикарбоновых кислот локализованы прямо в цитоплазме, а ферменты дыхательной цепи и фосфорилирования связаны с клеточной мембраной, с ее выпячиваниями, выступающими внутрь цитоплазмы, с так называемыми мезосомами (рис. 212). Надо отметить, что такие бактериальные мезосомы бывают связаны не только с процессами аэробного дыхания, но и у некоторых видов участвовать в делении клеток, в процессе распределения ДНК по новым клеткам, в образовании клеточной стенки и т.д. На плазматической мембране в мезосомах некоторых бактерий реализуются сопряженные процессы как окисления так и синтеза АТФ. В электронном микроскопе во фракциях плазматических мембран бактерий обнаружены сферические частицы, аналогичные тем, которые были найдены в митохондриях эукариотических клеток. Τᴀᴋᴎᴍ ᴏϬᴩᴀᴈᴏᴍ, у бактериальных клеток, способных к окислительному фосфорилированию, плазматическая мембрана выполняет роль, аналогичную внутренней мембране митохондрий эукариотических клеток.
Увеличение числа митохондрий. Митохондрии могут увеличивать свою численность особенно при делении клеток или при увеличении функциональной нагрузки клетки. Происходит постоянное обновление митохондрий. К примеру, в печени средняя продолжительность жизни митохондрий составляет около 10 дней.
Увеличение числа митохондрий происходит путем роста и деления предшествующих митохондрий. Это предположение было впервые высказано Альтманом (1893), описавшим митохондрии под термином “биобласты”. Удается наблюдать прижизненно деление, фрагментацию длинных митохондрий на более короткие путем перетяжки, что напоминает бинарный способ деления бактерий.
Реальное увеличение числа митохондрий путем деления установлено при изучении поведения митохондрий в живых клетках культуры ткани. В течение клеточного цикла митохондрии вырастают до нескольких мкм, а затем фрагментируются, делятся на более мелкие тельца.
Митохондрии могут сливаться друг с другом и размножаться по принципу: митохондрия от митохондрии.
Авторепродукция митохондрий. Двумембранные органеллы обладают полной системой авторепродукции. В митохондриях и пластидах существует ДНК, на которой синтезируются информационные, трансферные и рибосомные РНК и рибосомы, осуществляющие синтез митохондриальных и пластидных белков. При этом, эти системы, хотя и автономны, но ограничены по своим возможностям.
ДНК в митохондриях представляет собой циклические молекулы без гистонов и тем самым напоминают бактериальные хромосомы. Размер их составляет около 7 мкм, в одну циклическую молекулу митохондрий животных входит 16-19 тыс. нуклеотидных пар ДНК. У человека митохондриальная ДНК содержит 16,5 тыс. н.п., она полностью расшифрована. Найдено, что митохондральная ДНК различных объектов очень однородна, отличие их заключается лишь в величине интронов и нетранскрибируемых участков. Все митохондриальные ДНК представляют множественные копии, собранными в группы, кластеры. Так в одной митохондрии печени крысы может содержаться от 1 до 50 циклических молекул ДНК. Общее же количество митохондриальной ДНК на клетку составляет около одного процента. Синтез митохондриальных ДНК не связан с синтезом ДНК в ядре.
Так же как и у бактерий митохондральная ДНК собрана в отдельную зону – нуклеоид, его размер составляет около 0, 4 мкм в диаметре. В длинных митохондриях должна быть от 1 до 10 нуклеоидов. При делении длинной митохондрии от нее отделяется участок, содержащий нуклеоид (сходство с бинарным делением бактерий). Количество ДНК в отдельных нуклеоидах митохондрий может колебаться в 10 раз исходя из типа клеток.
В некоторых культурах в клетках от 6 до 60% митохондрий не имеют нуклеоида, что может объясняться тем, что деление этих органелл скорее связано с фрагментацией, а не с распределением нуклеоидов.
Как уже говорилось, митохондрии могут как делиться, так и сливаться друг с другом. При слиянии митохондрий друг с другом может происходить обмен их внутренними компонентами.
Важно подчеркнуть, что рРНК и рибосомы митохондрий и цитоплазмы резко отличны. В случае если в цитоплазме обнаруживаются 80s рибосомы, то рибосомы митохондрий растительных клеток принадлежат к 70s рибосомам (состоят из 30s и 50s субъединиц, содержат 16s и 23s РНК, характерные для прокариотических клеток), а в митохондриях клеток животных обнаружены более мелкие рибосомы (около 50s).
Рибосомные РНК митохондрий синтезируются на митохондриальных ДНК. В митоплазме на рибосомах идет синтез белков. Он прекращается, в отличие от синтеза на цитоплазматических рибосомах, при действии антибиотика хлорамфеникола, подавляющего синтез белка у бактерий.
На митохондриальном геноме синтезируются 22 транспортные РНК. Триплетный код митохондриальной синтетической системы отличен от такового, используемого в гиалоплазме. Несмотря на наличие казалось бы всех компонентов, необходимых для синтеза белков, небольшие молекулы митохондриальной ДНК не могут кодировать все митохондриальные белки, только лишь их небольшую часть. Так ДНК размером 15 тыс.н.п. может кодировать белки с суммарным молекулярным весом около 6х10 5 . В это же время суммарный молекулярный вес белков частицы полного дыхательного ансамбля митохондрии достигает величины около 2х10 6 . В случае если учесть, что кроме белков окислительного фосфорилирования в митохондрии входят ферменты цикла трикарбоновых кислот, ферменты синтеза ДНК и РНК, ферменты активации аминокислот и другие белки, то видно, что, для того чтобы кодировать эти многочисленные белки и рРНК и тРНК, количества генетической информации в короткой молекуле митохондриальной ДНК явно не хватает. Расшифровка нуклеотидной последовательности митохондриальной ДНК человека показала, что она кодирует всего лишь 2 рибосомные РНК, 22 трансферных РНК и всего 13 различных полипептидных цепей.
Сегодня доказано, что большая часть белков митохондрий находится под генетическим контролем со стороны клеточного ядра и синтезируется вне митохондрий. Большинство митохондриальных белков синтезируется на рибосомах в цитозоле. Эти белки имеют специальные сигнальные последовательности, которые узнаются рецепторами на внешней мембране митохондрий. Эти белки могут встраиваться в них (см. аналогию с мембраной пероксисом), а затем перемещаться на внутреннюю мембрану. Этот перенос происходит в точках контакта наружной и внутренней мембран, где такой транспорт отмечен (рис. 214). Большинство липидов митохондрий так же синтезируются в цитоплазме.
Все это говорит о эндосимбиотическом происхождении митохондрий, о том, что митохондрии представляют из себяорганизмы типа бактерий, находящиеся в симбиозе с эукариотический клеткой.
Хондриом. Совокупность всех митохондрий в одной клетке принято называть хондриомом. Она должна быть различной исходя из типа клеток. Во многих клетках хондриом состоит из разрозненных многочисленных митохондрий, равномерно расположенных по всей цитоплазме или локализуются группами в местах интенсивной траты АТФ. В обоих этих случаях митохондрии функционируют поодиночке, их кооперативная работа͵ возможно, координируется какими-то сигналами из цитоплазмы. Существует и совершенно иной тип хондриома, когда вместо мелких одиночных разрозненных митохондрий в клетке располагается одна гигантская разветвленная митохондрия (рис. 215в). Такие митохондрии встречаются у одноклеточных зеленых водорослей (к примеру у Chlorella). Οʜᴎ образуют сложную митохондриальную сеть или митохондриальный ретикулум (Reticulum miyochondriale). Согласно хемоосмотической теории биологический смысл появления такой гигантской разветвленной митохондриальной структуры, объединенной в одно целое своими внешними и внутренними мембранами состоит по сути в том, что в любой точке поверхности внутренней мембраны такой разветвленной митохондрии может идти синтез АТФ, который будет поступать в любую точку цитоплазмы, где в данном есть крайне важно сть.
В случае гигантских разветвленных митохондрий в любой ее точке может на внутренней мембране накопиться потенциал, достаточный для того, чтобы начался синтез АТФ. С этих позиций митохондриальный ретикулум представляет собой как бы электрический проводник, кабель, соединяющий отдаленные точки такой системы. Митохондриальный ретикулум оказался очень полезным не только для мелких подвижных клеток, таких как хлорелла, но и для более крупных структурных единиц таких как, к примеру, миофибриллы в скелетных мышцах.
Известно, что скелетные мышцы состоят из массы мышечных волокон, симпластов, содержащих множество ядер.
Размещено на реф.рф
Длина таких мышечных волокон достигает 40 мкм, при толщине 0,1 мкм - ϶ᴛᴏ гигантская структура, содержащая великое множество миофибрилл, все из которых сокращаются одновременно, синхронно. Важно заметить, что для сокращения к каждой единице сокращения, к миофибрилле, доставляется большое количество АТФ, которых обеспечивают митохондрии на уровне z-дисков. На продольных ультратонких срезах скелетных мышц в электронном микроскопе видны многочисленные округлые мелкие сечения митохондрий, располагающихся в соседстве с саркомерами (рис. 217). Мышечные митохондрии представляют из себяне мелкие шарики или палочки, а как бы паукообразные структуры, отростки которых ветвятся и простираются на большие расстояния, иногда через весь поперечник мышечного волокна. При этом разветвления митохондрий окружают каждую миофибриллу в мышечном волокне, снабжая их АТФ, крайне важно го для мышечного сокращения. Следовательно, в плоскости z-диска митохондрии представляют типичный митохондриальный ретикулум. Такой пласт или этаж митохондриального ретикулума повторяется дважды на каждый саркомер, а все мышечное волокно имеет тысячи поперечно расположенных “поэтажных” пластов митохондриального ретикулума. Между “этажами” вдоль миофибрилл располагаются нитчатые митохондрии, соединяющие эти митохондриальные пласты. Таким образом создана трехмерная картина митохондриального ретикулума, проходящего через весь объём мышечного волокна (рис. 218).
Далее было установлено, что между ответвлениями митохондриального ретикулума и нитевидными продольными митохондриями существуют специальные межмитохондриальные соединения или контакты (ММК). Οʜᴎ образованы плотно прилегающими наружными митохондриальными мембранами контактирующих митохондрий, межмембранное пространство и мембраны в этой зоне имеют повышенную электронную плотность (рис. 219). Через эти специальные образования происходит функциональное объединение соседних митохондрий и митохондриальных ретикулумов в единую, кооперативную энергетическую систему. Все миофибриллы в мышечном волокне сокращаются синхронно по всей их длине, следовательно, и поступление АТФ на любом участке этой сложной машины тоже должно происходить синхронно, а это может происходить лишь в том случае, в случае если огромное количество разветвленных митохондрий-проводников будет связано друг с другом с помощью контактов.
О том, что межмитоходриальные контакты (ММК) участвуют в энергетическом объединении митохондрий друг с другом удалось на кардиомиоцитах, клетках сердечных мышц.
Хондриом клеток сердечной мышцы не образует ветвящихся структур, а представлен множеством небольших вытянутых митохондрий, располагающихся без особого порядка между миофибриллами. При этом, все соседние митохондрии стыкуются друг с другом с помощью митохондриальных контактов такого же типа, как в скелетной мышце, только их число очень велико: в среднем на одну митохондрию приходится 2-3 ММК, которые связывают митохондрии в единую цепь, где каждым звеном такой цепи (Streptio mitochondriale) является отдельная митохондрия (рис. 220).
Оказалось, что межмитохондриальные контакты (ММК), как обязательная структура сердечных клеток обнаружены в кардиомиоцитах как желудочков, так и предсердий всех позвоночных животных: млекопитающих, птиц, пресмыкающихся, амфибий и костистых рыб. Более того ММК были обнаружены (но в меньшем числе) в клетках сердца некоторых насекомых и моллюсков.
Количество ММК в кардиомиоцитах изменяется исходя из функциональной нагрузки на сердце. Число ММК увеличивается при повышении физических нагрузок животных и, напротив - при падении нагрузки на сердечную мышцу происходит резкое сокращение числа ММК.
Митохондрии – строение и функции - понятие и виды. Классификация и особенности категории "Митохондрии – строение и функции" 2017, 2018.
Митохондрии
В клетках животных тканей митохондрии были обнаружены в 1882 г., а у растений - только в 1904 г. (в пыльниках кувшинки). Биологические функции удалось установить после выделения и очистки фракции методом фракционного центрифугирования. В их составе находится 70% белка и около 30% липидов, небольшое количество РНК и ДНК, витамины А, B 6 , В 12 , К, Е, фолиевая и пантотеновая кислоты, рибофлавин, различные ферменты. Митохондрии имеют двойную мембрану, Наружная изолирует органеллу от цитоплазмы, а внутренняя образует выросты кристы. Все пространство между мембранами заполнено матриксом (рис. 13).
Основная функция митохондрий - участие в клеточном дыхании. Роль митохондрий в дыхании была установлена в 1950-1951 годах. На наружных мембранах концентрируется сложная ферментная система цикла Кребса. При окислении субстратов дыхания освобождается энергия, которая тотчас же в процессе окислительного фосфорилирования, происходящего в кристах, аккумулируется в образующихся молекулах АДФ и главным образом АТФ. Энергия, запасенная в макроэргических соединениях, используется в дальнейшем для удовлетворения всех потребностей клетки.
Образование митохондрий в клетке происходит непрерывно из микротелец, чаще их возникновение связывают с дифференцировкой мембранных структур клетки. Они в клетке могут восстанавливаться путем их деления и почкования. Митохондрии не долговечны, продолжительность их жизни 5-10 дней.
Митохондрии – «силовые» станции клетки. В них концентрируется энергия, которая запасается в «аккумуляторах» энергии - молекулах АТФ, а не рассеивается в клетке. Нарушение структуры митохондрии ведет к нарушению процесса дыхания и в итоге к патологии организма.
Аппарат Гольджи. Аппарат Гольджи (синоним - диктиосомы) представляет собой стопки из 3-12 уплощенных, замкнутых, окруженных двойной мембраной дисков, называемых цистернами, от краев которых отшнуровываются многочисленные пузырьки (300-500). Ширина цистерн 6-90 А, толщина мембран - 60-70 А.
Аппарат Гольджи является центром синтеза, накопления и выделения полисахаридов, в частности целлюлозы, участвует в распределении и внутриклеточном транспорте белков, а также в образовании вакуолей и лизосом. В растительной клетке удалось проследить участие аппарата Гольджи в возникновении срединной пластинки и росте клеточной пекто-целлюлозной оболочки.
Аппарат Гольджи более всего развит в период активной жизни клетки. При ее старении он постепенно атрофируется, а затем исчезает.
Лизосомы. Лизосомы - довольно мелкие (около 0.5 мк в диаметре) округлые тельца. Они покрыты белково-липоидной мембраной. Содержимое лизосом многочисленные гидролитические ферменты, которые осуществляют функцию внутриклеточного переваривания (лизирования) макромолекул белка, нуклеиновых кислот, полисахаридов. Их основная функция переваривание отдельных участков протопласта клетки (автофагия - самопожирание). Этот процесс протекает за счет фагоцитоза или пиноцитоза. Биологическая роль этого процесса двоякая. Во-первых, защитная, поскольку при временном недостатке запасных продуктов клетка поддерживает жизнь за счет конституционных белков и др. веществ, а во-вторых происходит освобождение от избыточных или изношенных органелл (пластид, митохондрий и др.) Оболочка лизосомы препятствует выходу ферментов в цитоплазму, в противном случае она бы вся переваривалась этими ферментами.
В умершей клетке лизосомы разрушаются, ферменты оказываются в клетке и все ее содержимое переваривается. Остается только пекто-целлюлозная оболочка.
Лизосомы являются продуктами деятельности аппарата Гольджи, оторвавшимися от него пузырьками, в которых этот органоид аккумулировал переваривающие ферменты.
Сферосомы - округлые белково-липоидные тельца 0.3-0.4 мкм. По всей вероятности являются производными аппарата Гольджи или эндоплазматического ретикулума. По своей форме и величине напоминают лизосомы. Поскольку сферосомы содержат кислую фосфатазу, то они, вероятно, имеют отношение к лизосомам. Некоторые авторы считают, что сферосомы и лизосомы эквивалентны друг другу, но, скорее всего только по происхождению и форме. Есть предположение об их участии в синтезе жиров (А.Фрей-Висслинг).
Рибосомы - очень мелкие органоиды, диаметр их около 250А, По форме они почти шаровидные. Часть их прикреплена к наружным мембранам эндоплазматического ретикулума, часть их находится в свободном состоянии в цитоплазме. В клетке может содержаться до 5 млн рибосом. Рибосомы есть в хлоропластах и митохондриях, где они синтезируют часть белков, из которых построены эти органоиды, и ферменты, функционирующие в них.
Основная функция - синтез специфических белков согласно информации, поступающей из ядра. Их состав: белок и рибосомная рибонуклеиновая кислота (РНК) в равных соотношениях. Их структура малая и большая субъединицы, сформированные из рибонуклеотида.
Микротрубочки. Микротрубочки - своеобразные производные эндоплазматического ретикулума. Обнаружены во многих клетках. Само их название говорит об их форме - одна или две, расположенные параллельно, трубочки с полостью внутри. Внешний диаметр в пределах 250А. Стенки микротрубочек построены из белковых молекул. Из микротрубочек во время деления клетки образуются нити веретена.
Ядро
Ядро было обнаружено в растительной клетке Р. Броуном в 1831 году. Оно располагается в центре клетки или около клеточной оболочки, но со всех сторон окружено цитоплазмой. В большинстве случаев в клетке находится одно ядро, по несколько ядер находится в клетках некоторых водорослей, а также грибов. У зеленых водорослей неклеточной структуры насчитывается сотни ядер. Многоядерные клетки нечленистых млечников. Отсутствуют ядра в клетках бактерий и сине-зеленых водорослей.
Форма ядра чаще всего близка к форме шара или эллипса. Зависит от формы, возраста и функции клетки. В меристематической клетке ядро крупное, округлой формы и занимает 3/4 объема клетки. В паренхимных клетках эпидермы, имеющих крупную центральную вакуоль, ядро имеет чечевицеобразную форму и отодвинуто вместе с цитоплазмой к периферии клетки. Это признак специализированной, но уже стареющей клетки. Клетка, лишенная ядра, способна жить лишь короткое время. Безъядерные клетки ситовидных трубок живые клетки, но живут они недолго. Во всех других случаях безъядерные клетки являются мертвыми.
Ядро имеет двойную оболочку, через поры в которой содержимое
ядра (нуклеоплазма) может сообщаться с содержимым цитоплазмы. Мембраны оболочки ядра снабжены рибосомами и сообщаются с мембранами эндоплазматического ретикулума клетки. В нуклеоплазме располагается одно или два ядрышка и хромозомы. Нуклеоплазма представляет собой коллоидную систему золя, напоминающую по консистенции загустевшую желатину. В ядре, по данным отечественных биохимиков (Збарский И.Б. и др.), содержится четыре фракции белков: простых белков - глобулинов 20%, дезоксирибонуклеопротеидов - 70%, кислых белков - 6% и остаточных белков 4%. Они локализуются в следующих ядерных структурах: ДНК-протеиды (щелочные белки) - в хромозомах, РНК-протеиды (кислые белки) - в ядрышках, частично в хромозомах (в период синтеза информационной РНК) и в ядерной мембране. Глобулины составляют основу нуклеоплазмы. Остаточные белки (природа не уточнена) образуют ядерную мембрану.
Основная масса белков ядра - сложные щелочные белки дезоксирибонуклеопротеиды, в основе которых находится ДНК.
Молекула ДНК. Молекула ДНК – полинуклеотид и состоит из нуклеотидов. В состав нуклеотида входит три компонента: молекула сахара (дезоксирибоза), молекула азотистого основания и молекулы фосфорной кислоты. Дезоксирибоза соединена с азотистым основанием гликозидной, а с фосфорной кислотой - эфирной связью. В ДНК имеется в различных комбинациях всего 4 разновидности нуклеотидов, отличающихся друг от друга азотистыми основаниям. Два из них (аденин и гуанин) относятся к пуриновым азотистым соединениям, а цитозин и тимин - к пиримидиновым. Молекулы ДНК располагаются не в одной плоскости, а состоят из двух спирализованных нитей, т.е. две параллельно расположенные цепочки, закрученные одна вокруг другой, образуют одну молекулу ДНК. Они скреплены между собой с помощью водородной связи между азотистыми основаниями, причем пуриновые основания одной цепочки присоединяют пиримидиновые основания другой (рис.14). Структура и химизм молекулы ДНК была раскрыта английским (Крик) и американским (Уотсон) учеными и обнародована в 1953 г. Этот момент принято считать началом развития молекулярной генетики. Молекулярный вес ДНК – 4-8 млн. Количество нуклеотидов (различных вариантов) до 100 тысяч. Молекула ДНК очень стабильна, ее стабильность обеспечивается тем, что на всем протяжении она имеет одинаковую толщину - 20А (8А - ширина пиримидинового основания +12А - ширина пуринового основания). Если ввести в организм радиоактивный фосфор, то метка будет обнаруживаться во всех фосфоросодержащих соединениях, кроме ДНК (Леви, Сикевиц).
Молекулы ДНК являются носителями наследственности, т.к. в их структуре закодирована информация о синтезе специфических белков, определяющих свойства организма. Изменения могут возникнуть под действием мутагенных факторов (радиоактивное излучение, сильнодействующие, химические агенты -алкалоиды, спирты и т.д.).
Молекула РНК. Молекулы рибонуклеиновой кислоты (РНК) значительно меньше молекул ДНК. Это одиночные цепочки из нуклеотидов. Существует три вида РНК: рибосомная, самая длинная, образующая многочисленные петли, информационная (матричная) и траспортная, самая короткая. Рибосомная РНК локализуется в рибосомах эндоплазматической сети и составляет 85% всей РНК клетки.
Информационная РНК по своей структуре напоминает листочек клевера. Ее количество - 5% от суммы всей РНК в клетке. Она синтезируется в ядрышках. Ее сборка происходит в хромозомах в период интерфазы. Ее основная функция - перенос информации от ДНК к рибосомам, где происходит синтез белка.
Транспортная РНК, как установлено сейчас, это целое семейство соединений, родственных по структуре и биологической функции. Каждая живая клетка по приблизительной оценке содержит 40-50 индивидуальных транспортных РНК и их общее число в природе, если учесть видовые различия, огромно. (Акад.В.Энгельгардт). Транспортными они называются потому, что их молекулы заняты транспортным обслуживанием внутриклеточного процесса синтеза белка. Соединяясь со свободными аминокислотами, они доставляют их к рибосомам в строящуюся белковую цепь. Это самые маленькие молекулы РНК, состоят в среднем из 80 нуклеотидов. Локализуются в матриксе цитоплазмы и составляют около 10% клеточной РНК
В составе РНК содержится четыре азотистых основания, но в отличие от ДНК в молекуле РНК вместо тимина находится урацил.
Структура хромозом. Хромозомы впервые были обнаружены в конце 19 века классиками цитологии Флемингом и Страсбургером (1882, 1884), а русский исследователь клетки И.Д. Чистяков их обнаружил в 1874 году.
Основной структурный элемент хромозом - ядро. Они имеют различную форму. Это либо прямые, либо изогнутые палочки, овальные тельца, шарики, размеры которых варьируют.
В зависимости от места расположения центромеры различают прямые, равноплечие и неравноплечие хромозомы. Внутренняя структура хромозом представлена на рис. 15, 16. Следует отметить, что дезоксирибонуклеопротеид является мономером хромозомы.
В хромозоме дезоксирибонуклеопротеидов 90-92%, из них 45% -ДНК и 55% - белка (гистона). В небольшом количестве в хромозоме представлена и РНК (информационная).
У хромозомы четко выражена и поперечная структура - наличие утолщенных участков - дисков, которые еще в 1909г. были названы генами. Этот термин был предложен датским ученым Иогансеном. В 1911 г. американский ученый Морган доказал, что гены являются основными наследственными единицами и распределяются они в хромозомах в линейном порядке и, поэтому хромозома имеет качественно различные участки. В 1934 г. американский ученый Пайнтер доказал прерывистость морфологического строения хромозом и наличие в хромозомах дисков, а диски - это места скопления ДНК. Это послужило началом создания хромосомных карт, на которых указывалось место (локус) расположения гена, определяющего тот или иной признак организма. Ген - это участок двойной спирали ДНК, на котором содержится информация о структуре одного белка. Это участок молекулы ДНК, определяющий синтез одной молекулы белка. ДНК непосредственного участия в синтезе белка не принимает. В ней только содержится и хранится информация о структуре белка.
Структура ДНК, состоящая из нескольких тысяч последовательно расположенных 4-х нуклеотидов, представляет собой код наследственности.
Код наследственности. Синтез белка. Первое сообщение по коду ДНК сделал американский биохимик Ниренберг в 1961 г. в Москве на международном биохимическом конгрессе. Сущность кода ДНК состоит в следующем. Каждой аминокислоте соответствует участок цепи ДНК из трех рядом расположенных нуклеотидов (триплет). Так, например участок, состоящий из Т-Т-Т (триплет из 3-х тиминсодержащих нуклеотидов) соответствует аминокислоте лизину, триплет А (аденин) - Ц (цитозин) - А (аденин)- цистеину и т.д. Допустим, что ген представлен цепочкой нуклеотидов, расположенных в следующем порядке: А-Ц-А-Т-Т-Т-А-А-Ц-Ц-А-А-Г-Г-Г. Разбив этот ряд на триплеты, мы сразу расшифруем, какие аминокислоты и в каком порядке будут располагаться в синтезируемом белке.
Число возможных сочетаний из 4-х имеющихся нуклеотидов по три равно 4×64. Исходя из этих соотношений, числа различных триплетов с избытком хватит для обеспечения информации по синтезу многочисленных белков, определяющих и структуру и функции организма. Для синтеза белка в рибосомы направляется точная копия этой информации в виде информационной РНК. В расшифровке и синтезе, кроме и-РНК, участвует большое число молекул различных транспортных рибонуклеиновых кислот (т-РНК), рибосомы и ряд ферментов. Каждая из 20 аминокислот связывается с Т-РНК - молекула с молекулой. Каждой из 20 аминокислот соответствует своя т-РНК. У т-РНК имеются химические группы, способные «узнавать» свою аминокислоту, выбирая именно ее из наличных аминокислот. Происходит это с помощью специальных ферментов. Узнав свою аминокислоту, т-РНК вступает с ней в соединение. К началу цепочки (молекулы) и-РНК присоединяется рибосома, которая, продвигаясь по и-РНК, соединяет друг с другом в полипептидную цепочку именно те аминокислоты, порядок которых зашифрован нуклеотидной последовательностью данной И-РНК. В результате образуется молекула белка, состав которого закодирован в одном из генов.
Ядрышки - неотъемлемая структурная часть ядра. Это сферические тельца. Они очень изменчивы, меняют свою форму и структуру, появляются и исчезают. Их бывает одно, два. Для каждого растения определенное число. Ядрышки исчезают, когда клетка готовится к делению, а затем появляются вновь; они, по-видимому, участвуют в синтезе рибонуклеиновых кислот. Если ядрышко разрушить сфокусированным пучком рентгеновских или ультрафиолетовых лучей, то клеточное деление подавляется.
Роль ядра в жизни клетки. Ядро служит контролирующим центром клетки- оно направляет клеточную активность и содержит носителей наследственности (гены), определяющие признаки данного организма. Роль ядра можно выявить, если с помощью микрохирургических приемов удалить его из клетки и наблюдать последствие этого. Ряд опытов, доказывающих важную роль в регуляции клеточного роста, провел Геммерлинг на одноклеточной зеленой водоросли Acetobularia. Эта морская водоросль достигает высоты 5 см, внешне напоминает гриб, имеет подобие "корней" и "ножки". Вверху заканчивается большой дисковидной "шляпкой". Клетка этой водоросли имеет одно ядро, располагающееся в базальной части клетки.
Гаммерлинг установил, что если перерезать ножку, то нижняя часть продолжает жить и полностью регенерируют шляпку после операции. Верхняя же часть, лишенная ядра, выживает в течение некоторого времени, но, в конце концов, погибает, не будучи в состоянии восстановить нижнюю часть. Следовательно, ацетобулярии ядро необходимо для метаболических реакций, лежащих в основе роста.
Ядро способствует образованию клеточной оболочки. Это можно проиллюстрировать экспериментами с водорослью Voucheria и Spyrogyra. Выпуская из перерезанных нитей в воду содержимое клеток, мы можем получить комочки цитоплазмы с одним, с несколькими ядрами и без ядер. В первых двух случаях клеточная оболочка формировалась нормально. В случае отсутствия ядра оболочка не образовывалась.
В опытах И.И.Герасимова (1890г.) со спирогирой было установлено, что клетки с двойным ядром удваивают длину и толщину хлоропласта. В безъядерных клетках продолжается процесс фотосинтеза, образуется ассимиляционный крахмал, но при этом затухает процесс его гидролиза, что объясняется отсутствием гидролитических ферментов, которые могут быть синтезированы в рибосомах лишь согласно информации ДНК ядра. Жизнь протопласта без ядра неполноценна и недолговечна. В экспериментах И.И. Герасимова безъядерные клетки спирогиры жили 42 дня и погибали. Одна из важнейших функций ядра состоит в снабжении цитоплазмы рибонуклеиновой кислотой, необходимой для синтеза белка в клетке. Удаление ядра из клетки ведет к постепенному падению содержания РНК в цитоплазме и замедлению синтеза белка в ней.
Наиболее важна роль ядра в передаче признаков от клетки клетке, от организма к организму и осуществляет это в процессе деления ядра и клетки в целом.
Клеточное деление. Размножаются клетки делением. При этом из одной клетки образуется две дочерних с тем же набором наследственного материала, заключенного в хромозомах, что и материнская клетка. В соматических клетках хромозомы представлены двумя, так называемыми гомологическими хромозомами, в которых заложены аллельные гены (носители противоположных признаков, например, белый и красный цвет лепестков гороха и т.д.), признаков двух родительских пар. В связи с этим в соматических клетках тела растения всегда удвоенный набор хромозом, обозначаемый 2п. Хромозомы обладают выраженной индивидуальностью. Количество и качество хромозом - характерный признак каждого вида. Так, в клетках земляники диплоидный набор хромозом равен 14, (2n), яблони -34, топинамбура - 102 и т.д.
Митоз (кариокинез) – деление соматических клеток был впервые описан Э. Руссовым(1872г.) и И.Д.Чистяковым (1874). Его сущность заключается в том, что из материнской клетки путем деления образуется две дочерние клетки с тем же набором хромозом.Клеточный цикл слагается из интерфазы и собственно митоза. Методом микроавторадиографии установлено, что самой длительной и сложной является интерфаза - период "покоящегося" ядра, т.к. в этот период происходит удвоение ядерного материала. Интерфаза делится на три фазы:
Q1 - пресинтетическая (ее длительность 4-6 часов);
S - синтетическая (10-20 часов);
Q2 - постсинтетическая (2-5 часов).
Во время Q1 фазы идет подготовка к редупликации ДНК. А в S-фазу происходит редупликация ДНК, клетка удваивает запас ДНК. В Q2-фазу формируются ферменты и структуры, необходимые для запуска митоза. Таким образом, в интерфазе молекулы ДНК в хромозомах расщепляются на две одинаковые нити, происходит сборка на их матрице информационных РНК. Последняя уносит информацию о структуре специфических белков в цитоплазму, а в ядре каждая из нитей ДНК достраивает недостающую половинку своей молекулы. В этом процессе удвоения (редупликация) проявляется уникальная особенность ДНК, состоящая в способности ДНК точно воспроизводить саму себя. Образовавшиеся дочерние молекулы ДНК автоматически получаются точными копиями родительской молекулы, т.к. при редупликации к каждой половинке присоединяются комплементарные (А-Т; Г-Ц; и т.д.) основания из окружающей среды.
В профазу митотического деления удвоенные хромозомы становятся заметными. В метафазе все они располагаются в экваториальной зоне, располагаясь в один ряд. Образуются нити веретена (из микротрубочек, соединяющихся друг с другом). Оболочка ядра и ядрышко исчезают. Утолщенные хромозомы расщепляются вдоль на две дочерние хромозомы. В этом заключается суть митоза. Он обеспечивает точное распределение удвоенных молекул ДНК между дочерними клетками. Тем самым обеспечивает и передачу зашифрованной в ДНК наследственной информации.
В анафазе дочерние хромозомы начинают отходить к противоположным полюсам. В центре появляются первые фрагменты клеточной оболочки (фрагмобласт).
В телофазе происходит оформление ядер в дочерних клетках. Содержимое материнской клетки (органеллы) распределяется между образующимися дочерними. Полностью формируется клеточная оболочка. На этом заканчивается цитокинез (рис.17).
Мейоз - редукционное деление был обнаружен и описан в 90-х годах прошлого столетия В.И.Беляевым. Сущность деления заключается в том, что из соматической клетки, содержащей 2п (двойной, диплоидный) набор хромозом, образуется четыре гаплоидных клетки, с"n", половинным набором хромозом. Этот тип деления является сложным и состоит из двух этапов. Первый - редукция хромозом. Удвоенные хромозомы располагаются в экваториальной зоне попарно (две параллельно расположенные гомологичные хромозомы). В этот момент может происходить коньюгация (сцепление) хромозом, кроссинговер (перекрест) и в результате - обмен участками хромозом. В результате этого часть генов отцовских хромозом переходит в состав материнских хромозом и наоборот. Внешний вид тех и других хромозом в результате этого не меняется, но их качественный состав становится иным. Отцовская и материнская наследственности перераспределяются и смешиваются.
В анафазе мейоза гомологичные хромозомы с помощью нитей веретена расходятся по полюсам, на которых после небольшого периода покоя (исчезают нити, но перегородка между новыми ядрами не формируется) начинается процесс митоза - метафаза, при которой все хромозомы располагаются в одной плоскости и происходит их продольное расщепление на дочерние хромозомы. При анафазе митоза с помощью веретена они расходятся по полюсам, где и формируется четыре ядра и в итоге - четыре гаплоидные клетки. В клетках некоторых тканей при их развитии под влиянием некоторых факторов происходит незавершенный митоз и количество хромозом в ядрах удваивается за счет того, что не расходятся по полюсам. В результате таких нарушений естественного или искусственного характера возникают организмы тетраплоиды и полиплоиды. С помощью мейоза формируются половые клетки - гаметы, а также споры, элементы полового и бесполого размножения растений (рис.18).
Амитоз - прямое деление ядра. При амитозе веретено деления не образуется и оболочка ядра не распадается, как при митозе. Раньше амитоз рассматривался как примитивная форма деления. Сейчас установлено, что он связан с деградацией организма. Представляет собой упрощенный вариант более сложного деления ядра. Амитоз встречается в клетках и тканях нуцеллуса, эндосперма, паренхиме клубней, черешков листьев и т.д.
Митохондрии – преобразователи энергии и её поставщики для обеспечения клеточных функций – занимают значительную часть цитоплазмы клеток и сосредоточены в местах высокого потребления АТФ (например, в эпителии канальцев почки они располагаются вблизи плазматической мембраны (обеспечение реабсорбции), а в нейронах – в синапсах (обеспечение электрогенеза и секреции). Количество митохондрий в клетке измеряется сотнями. Митохондрии имеют собственный геном. Органелла функционирует в среднем 10 суток, обновление митохондрий происходит путем их деления.
Морфология митохондрии
Митохондрии чаще имеют форму цилиндра диаметром 0,2-1 мкм и длиной до 7 мкм (в среднем около 2 мкм). У митохондрий две мембраны – наружная и внутренняя; последняя образует кристы. Между наружной и внутренней мембранами находится межмембранное пространство. Внемембранный объем митохондрии – матрикс.
∙ Наружная мембрана проницаема для многих мелких молекул.
∙ Межмембранное пространство. Здесь накапливаются ионы Н + , выкачиваемые из матрикса, что создает протонный градиент концентрации по обе стороны внутренней мембраны.
∙ Внутренняя мембрана избирательно проницаема; содержит транспортные системы для переноса веществ (АТФ, АДФ, Р 1 , пирувата, сукцината, α-кетоглурата, малата, цитрата, цитидинтрифосфата, ГТФ, дифосфатов) в обоих направлениях и комплексы цепи переноса электронов, связанные с ферментами окислительного фосфорилирования, а также с сукцинатдегидрогеназой (СДГ).
∙ Матрикс. В матриксе присутствуют все ферменты цикла Кребса (кроме СДГ), ферменты β-окисления жирных кислот и некоторые ферменты других систем. В матриксе находятся гранулы с Mg 2+ и Ca 2+ .
∙ Цитохимические маркёры митохондрий – цитохромоксидаза и СДГ.
Функции митохондрий
Митохондрии выполняют в клетке множество функций: окисление в цикле Кребса, транспорт электронов, хемиосмотическое сопряжение, фосфорилирование АДФ, сопряжение окисления и фосфорилирования, функцию контроля внутриклеточной концентрации кальция, синтез белков, образование тепла. Велика роль митохондрий в программированной (регулируемой) гибели клеток.
∙ Теплорепродукция. Естественный механизм разобщения окислительного фосфорилирования функционирует в клетках бурого жира. В этих клетках митохондрии имеют атипичную структуру (уменьшен их объем, увеличена плотность матрикса, расширены межмембранные пространства) – конденсированные митохондрии. Такие митохондрии могут усиленно захватывать воду и набухать в ответ на тироксин, увеличение концентрации Ca 2+ в цитозоле, при этом усиливается разобщение окислительного фосфорилирования, и происходит выделение тепла. Эти процессы обеспечивает специальный разобщающий белок термогенин. Норадреналин из симпатического отдела вегетативной нервной системы усиливает экспрессию разобщающего белка и стимулирует теплопродукцию.
∙ Апоптоз. Митохондрии играют важную роль в регулируемой (программированной) гибели клеток – апоптозе, выделяя их в цитозоль факторы, повышающие вероятность гибели клетки. Одним из них является цитохром С – белок, переносящий электроны между белковыми комплексами во внутренней мембране митохондрий. Выделяясь из митохондрий, цитохром С включается в состав апоптосомы, активирующей каспазы (представители семейства киллерных протеаз).